

Myosin-1c interacts with hair-cell receptors through its calmodulin-binding IQ domains
- PMID: 11923413
- PMCID: PMC6758312
- DOI: 10.1523/JNEUROSCI.22-07-02487.2002
Myosin-1c interacts with hair-cell receptors through its calmodulin-binding IQ domains
Abstract
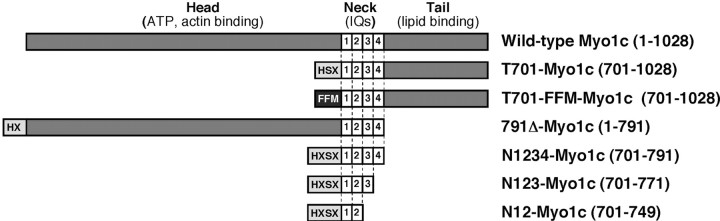
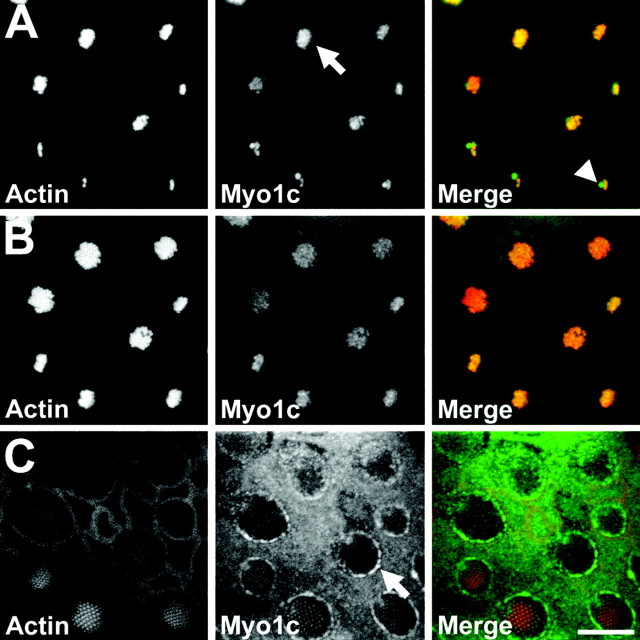
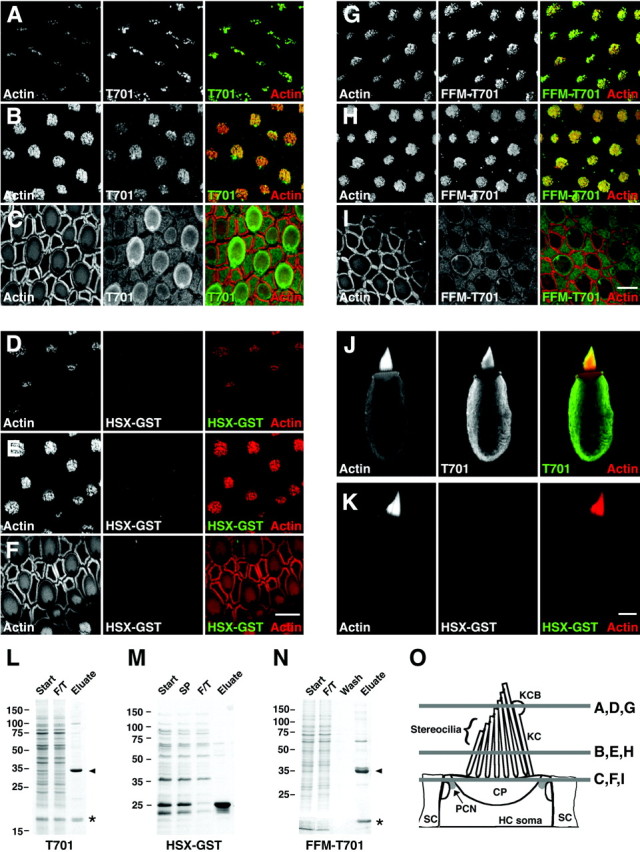
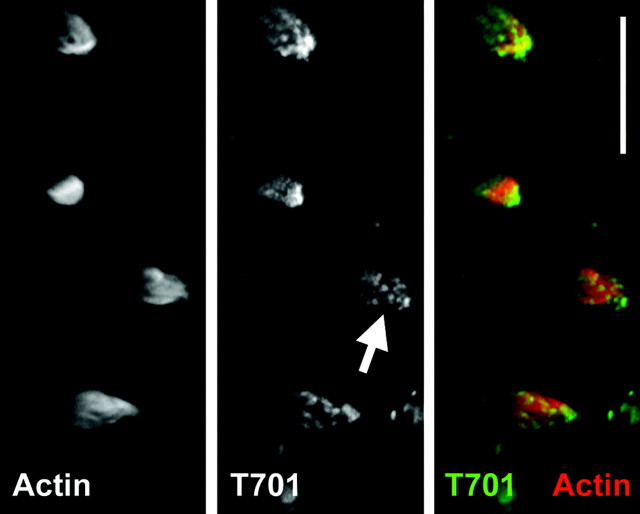
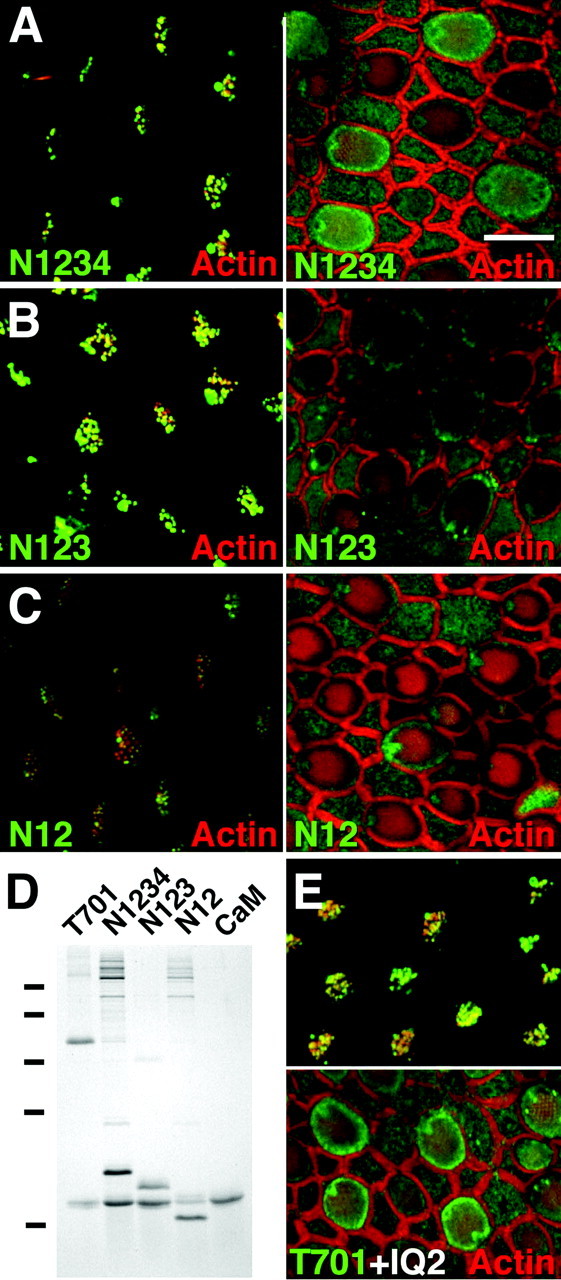
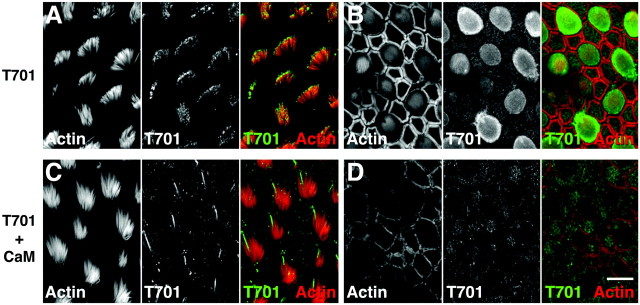
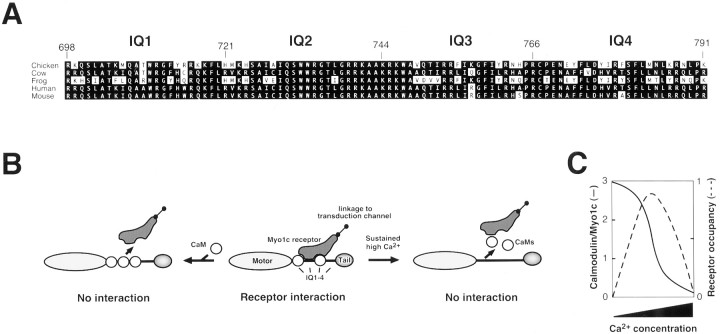
Myosin-1c plays an essential role in adaptation of hair-cell mechanoelectrical transduction. To mediate adaptation, myosin-1c must interact directly or indirectly with other components of the transduction apparatus, including the mechanically gated transduction channel. As a first step toward identifying myosin-1c receptors, we used recombinant myosin-1c fragments to identify specific binding sites in hair cells and to biochemically characterize their interaction with myosin-1c. Myosin-1c fragments bound to tips of hair-cell stereocilia, the location of transduction and adaptation. Surprisingly, this interaction did not depend on the C-terminal tail of myosin-1c, proposed previously to be the receptor-binding site of the molecule. Instead, the interaction of myosin-1c with stereociliary receptors depended on its calmodulin-binding IQ domains. This interaction was blocked by calmodulin, which probably bound to a previously unoccupied IQ domain of myosin-1c. The calcium-sensitive binding of calmodulin to myosin-1c may therefore modulate the interaction of the adaptation motor with other components of the transduction apparatus.
Figures
References
-
- Aruffo A, Stamenkovic I, Melnick M, Underhill CB, Seed B. CD44 is the principal cell surface receptor for hyaluronate. Cell. 1990;61:1303–1313. - PubMed
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials