

Cdk5/p35 regulates neurotransmitter release through phosphorylation and downregulation of P/Q-type voltage-dependent calcium channel activity
- PMID: 11923424
- PMCID: PMC6758295
- DOI: 10.1523/JNEUROSCI.22-07-02590.2002
Cdk5/p35 regulates neurotransmitter release through phosphorylation and downregulation of P/Q-type voltage-dependent calcium channel activity
Abstract
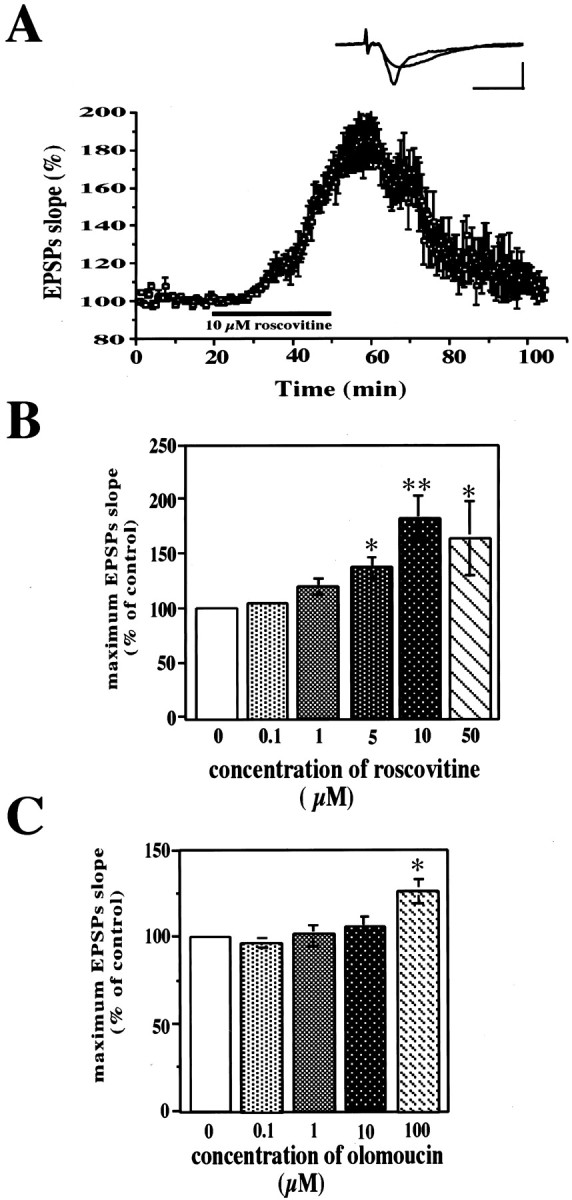
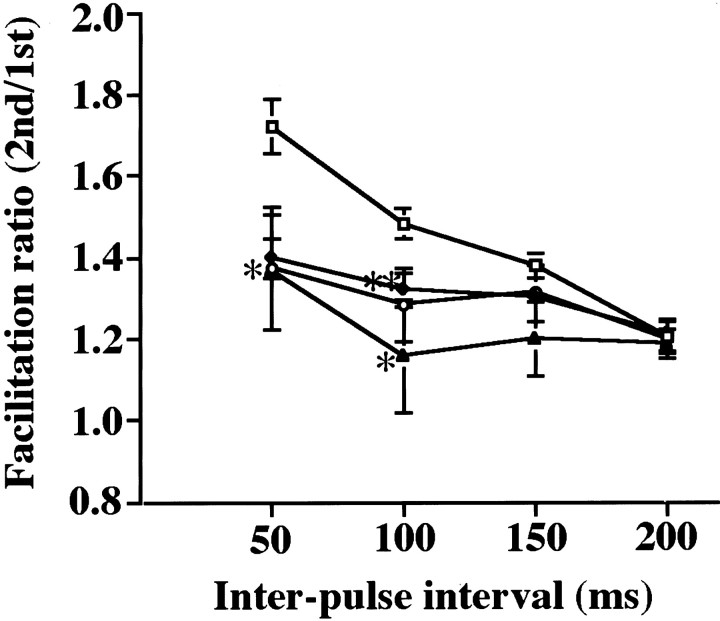
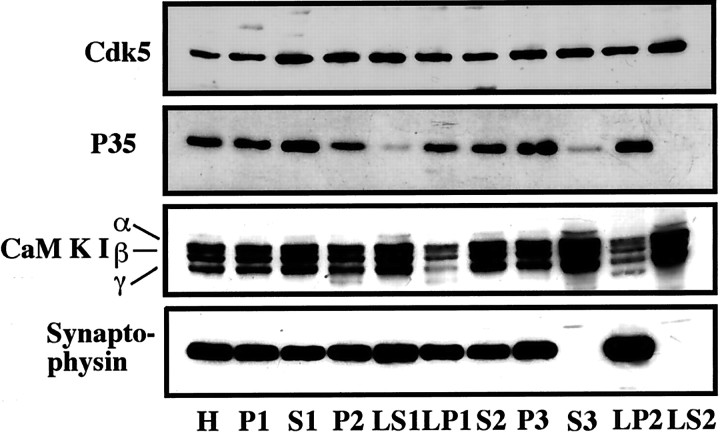
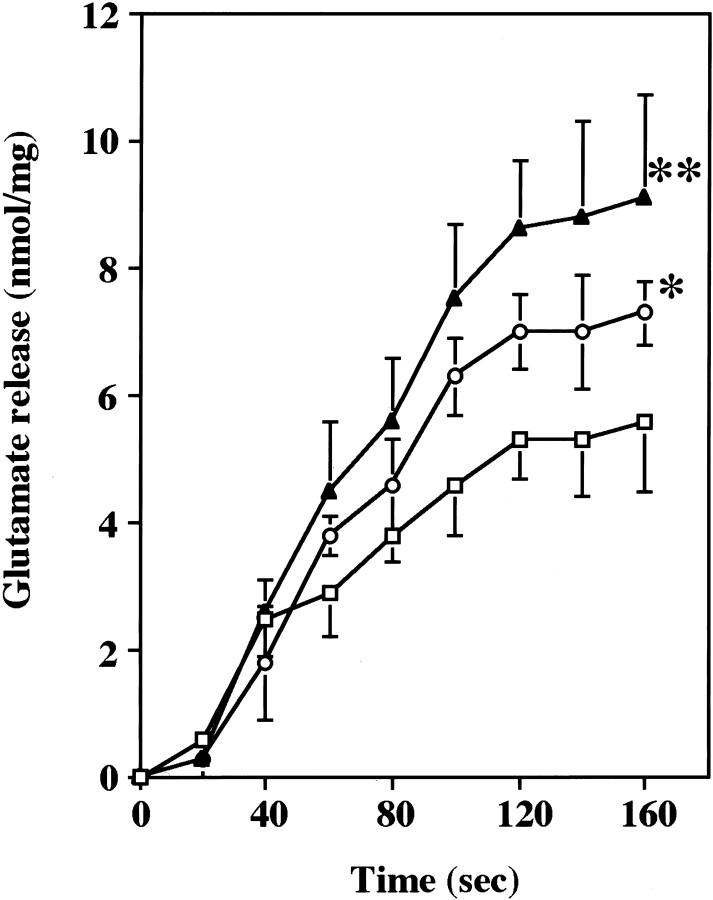
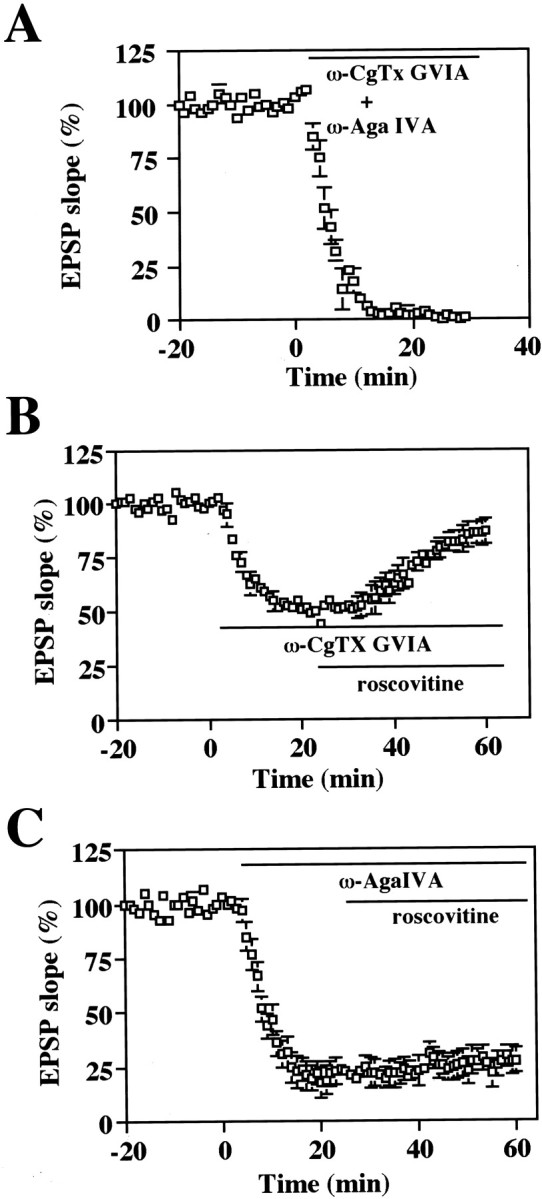
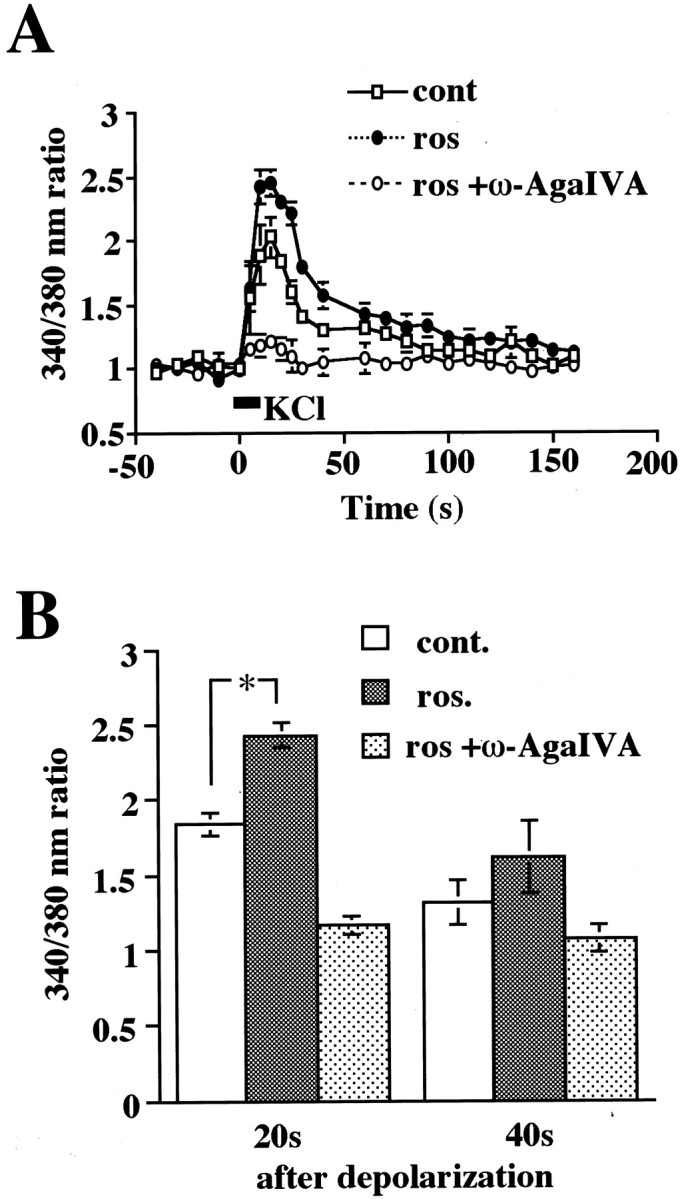
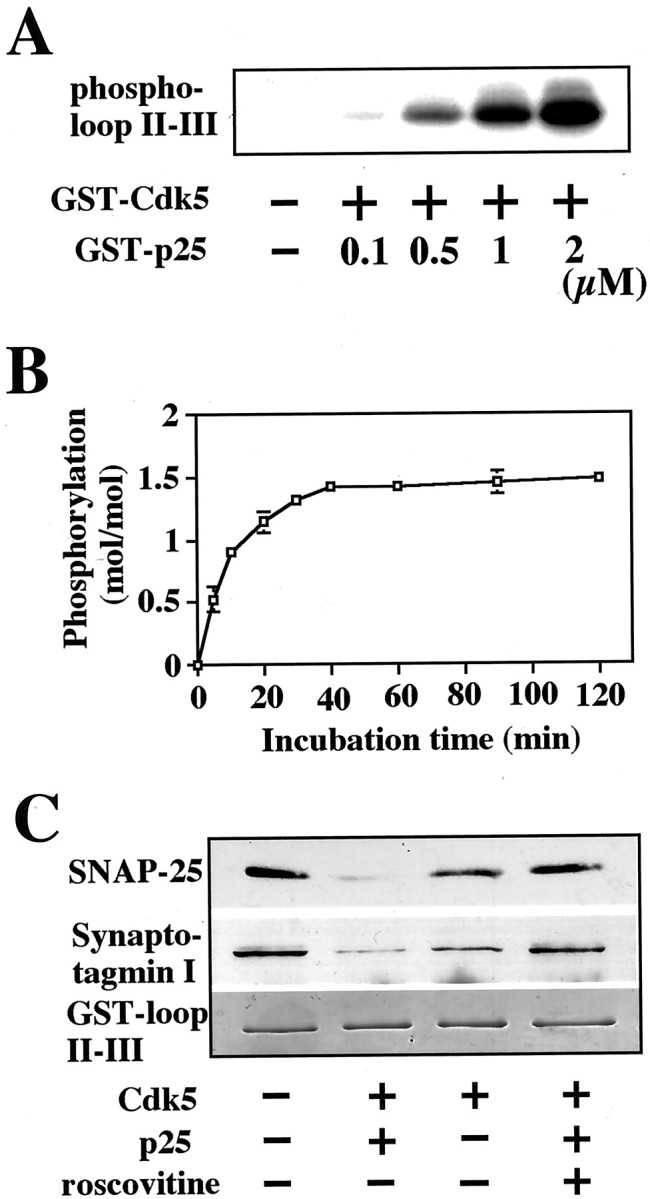
Cyclin-dependent kinase 5 (Cdk5) is a proline-directed serine/threonine kinase with close structural homology to the mitotic Cdks. The complex of Cdk5 and p35, the neuron-specific regulatory subunit of Cdk5, plays important roles in brain development, such as neuronal migration and neurite outgrowth. Moreover, Cdk5 is thought to be involved in the promotion of neurodegeneration in Alzheimer's disease. Cdk5 is abundant in mature neurons; however, its physiological functions in the adult brain are unknown. Here we show that Cdk5/p35 regulates neurotransmitter release in the presynaptic terminal. Both Cdk5 and p35 were abundant in the synaptosomes. Roscovitine, a specific inhibitor of Cdk5 in neurons, induced neurotransmitter release from the synaptosomes in response to membrane depolarization and enhanced the EPSP slopes in rat hippocampal slices. The electrophysiological study using each specific inhibitor of the voltage-dependent calcium channels (VDCCs) and calcium imaging revealed that roscovitine enhanced Ca2+ influx from the P/Q-type VDCC. Moreover, Cdk5/p25 phosphorylated the intracellular loop connecting domains II and III (L(II-III)) between amino acid residues 724 and 981 of isoforms cloned from rat brain of the alpha1A subunit of P/Q-type Ca2+ channels. The phosphorylation inhibited the interaction of L(II-III) with SNAP-25 and synaptotagmin I, which were plasma membrane soluble N-ethylmaleimide-sensitive factor attachment protein (SNAP) receptor (SNARE) proteins and were required for efficient neurotransmitter release. These results strongly suggest that Cdk5/p35 inhibits neurotransmitter release through the phosphorylation of P/Q-type VDCC and downregulation of the channel activity.
Figures
References
-
- Bibb JA, Snyder GL, Nishi A, Yan Z, Meijer L, Fienberg AA, Tsai LH, Kwon YT, Girault JA, Czernik AJ, Huganir RL, Hemmings HC, Jr, Nairn AC, Greengard P. Phosphorylation of DARPP-32 by Cdk5 modulates dopamine signalling in neurons. Nature. 1999;402:669–671. - PubMed
-
- Catterall WA. Structure and function of neuronal Ca2+ channels and their role in neurotransmitter release. Cell Calcium. 1998;24:307–323. - PubMed
-
- Chae T, Kwon YT, Bronson R, Dikkes P, Li E, Tsai LH. Mice lacking p35, a neuronal specific activator of Cdk5, display cortical lamination defects, seizures, and adult lethality. Neuron. 1997;18:29–42. - PubMed
-
- Dunlap K, Luebke JI, Turner TJ. Exocytotic Ca2+ channels in mammalian central neurons. Trends Neurosci. 1995;18:89–98. - PubMed
-
- Floyd SR, Porro EB, Slepnev VI, Ochoa GC, Tsai LH, De Camilli P. Amphiphysin binds the cdk5 regulatory subunit p35 and is phosphorylated by cdk5 and cdc2. J Biol Chem. 2001;276:8104–8110. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous