

Development of response timing and direction selectivity in cat visual thalamus and cortex
- PMID: 11923459
- PMCID: PMC6758333
- DOI: 10.1523/JNEUROSCI.22-07-02945.2002
Development of response timing and direction selectivity in cat visual thalamus and cortex
Abstract
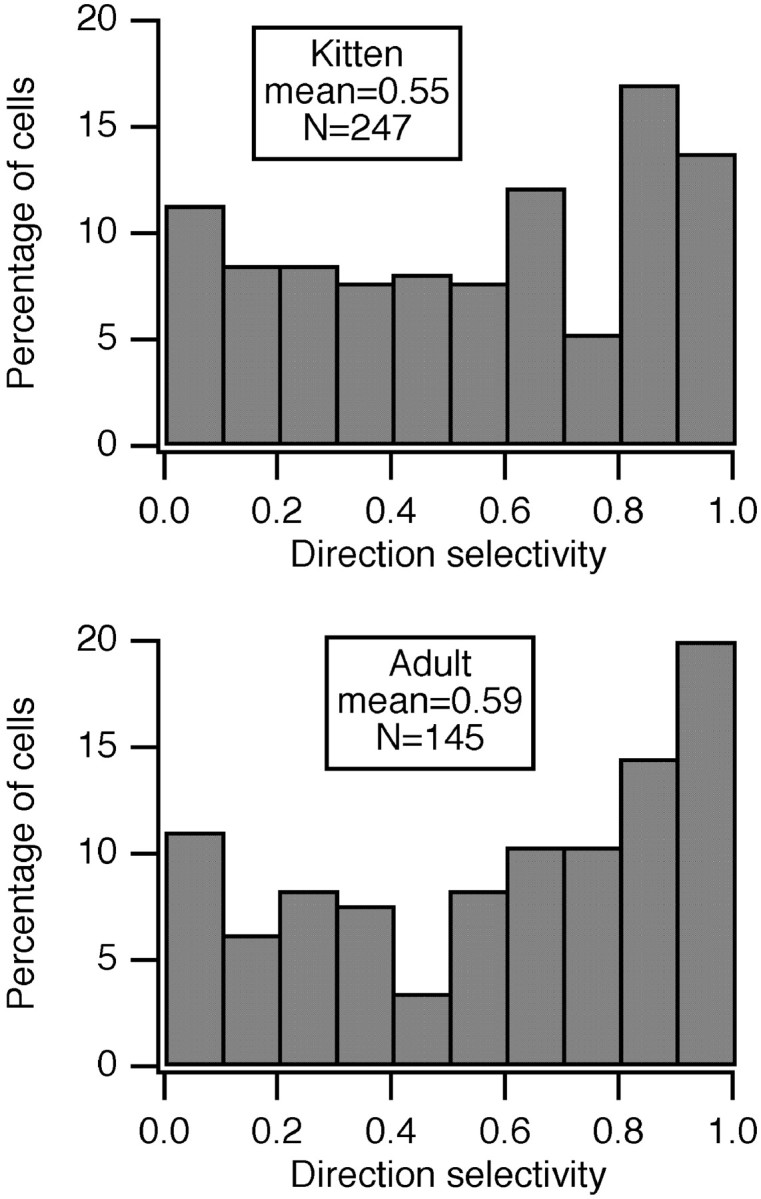
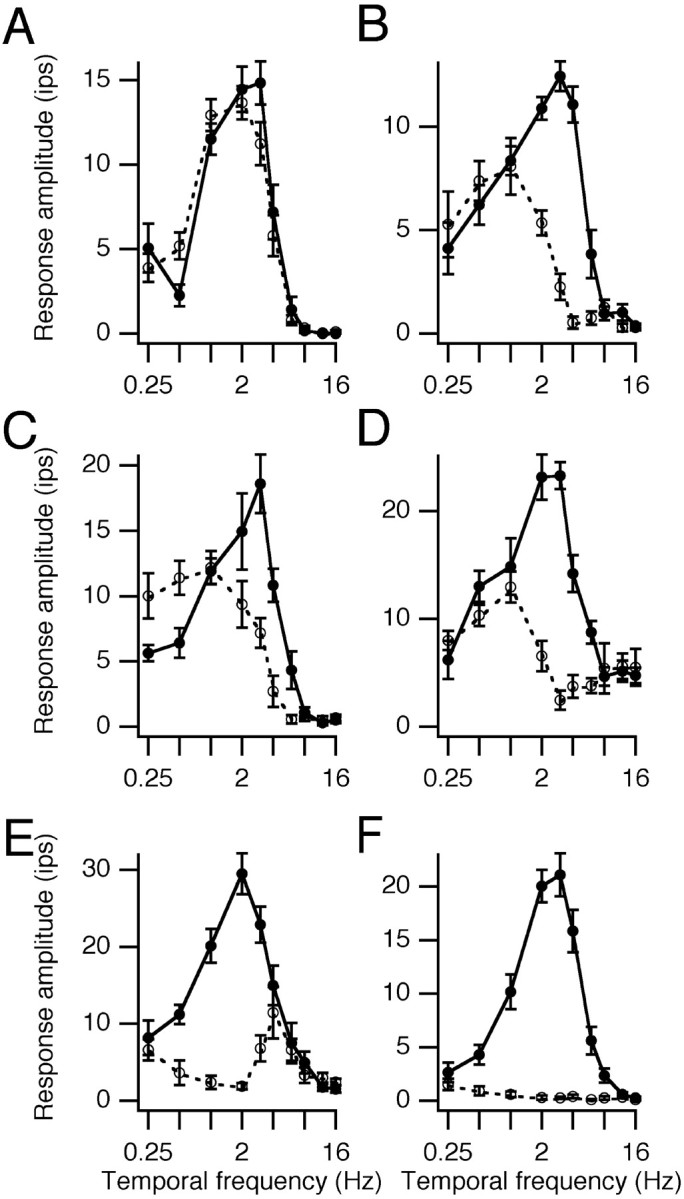
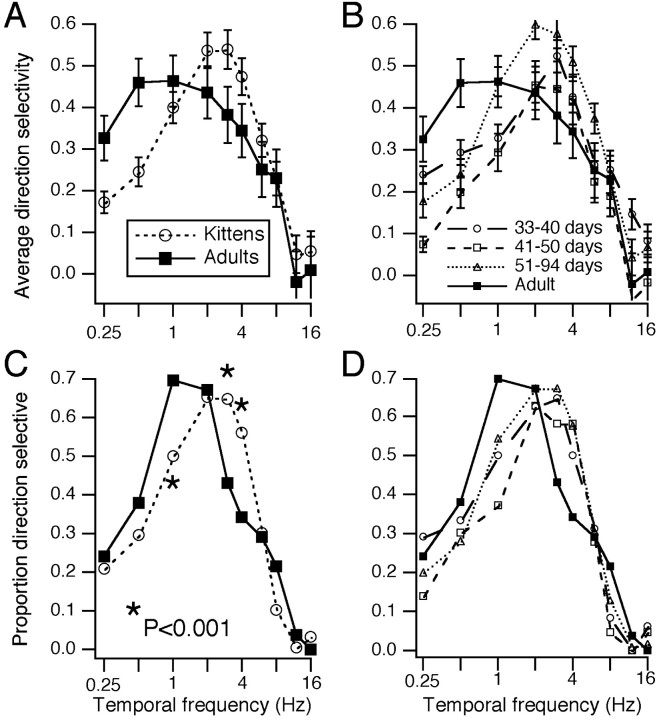
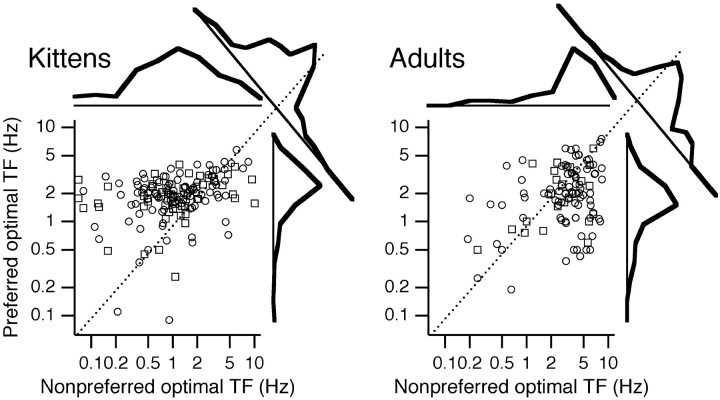
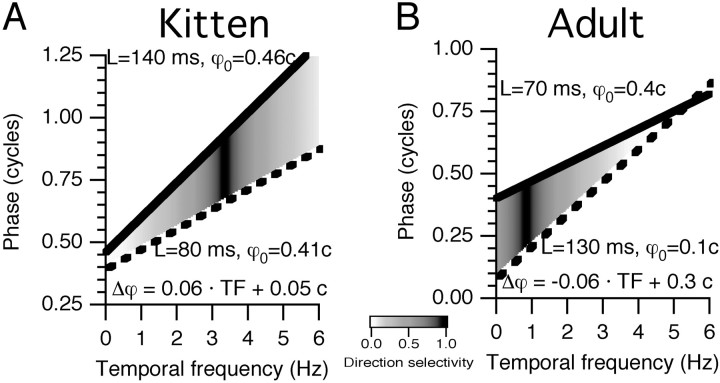
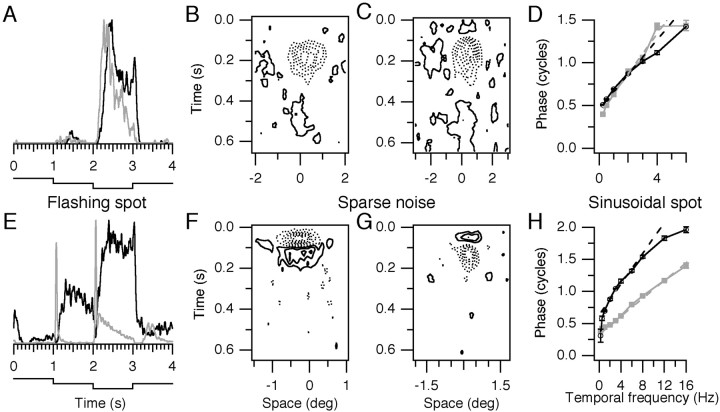
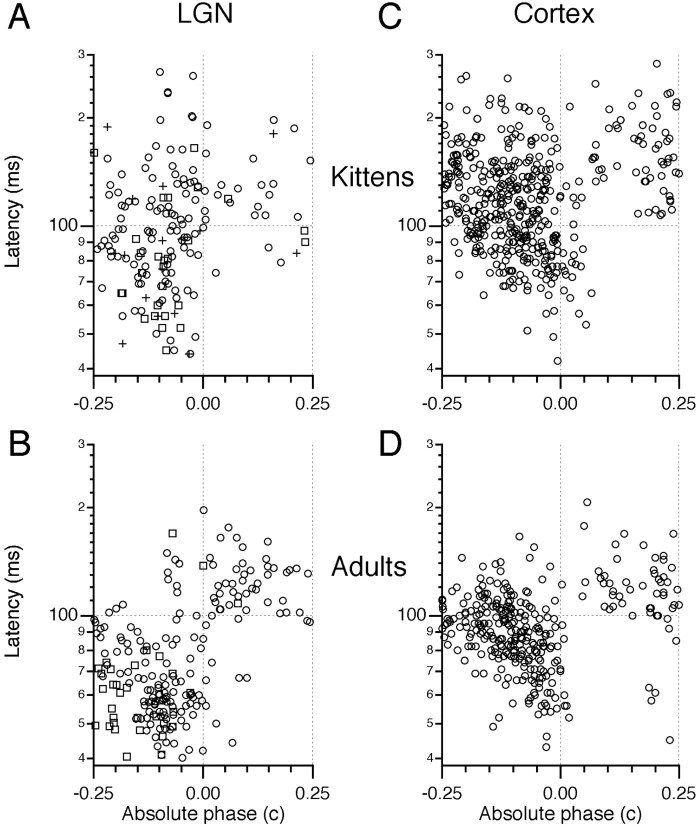
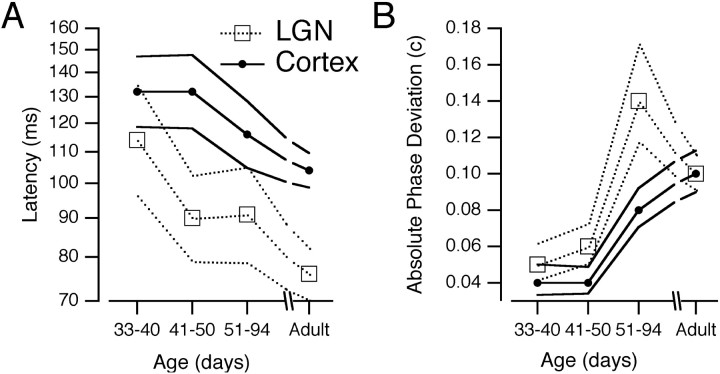
Single-unit recordings were made in the dorsal lateral geniculate nucleus (LGN) and visual cortex of kittens that were 4-13 weeks of age. Responses to visual stimuli were analyzed to determine the relationship between two related facets of the behaviors of the cells: direction selectivity (DS) and timing. DS depends on timing differences within the receptive field. Cortical DS was present at all ages, but its temporal frequency tuning changed. In kittens, DS was more common at high (approximately 4 Hz) than low ( approximately 1 Hz) temporal frequencies. This is in contrast to adults, in which DS is tuned to low frequencies, more common at 1 Hz than at 4 Hz (Saul and Humphrey, 1992a). In adult cats, the LGN provides the cortex with a wide range of timings that are also observable in cortical receptive fields (Saul and Humphrey, 1990, 1992b; Alonso et al., 2001). In kittens, LGN and cortical timing were immature. Most cells showed long-latency sustained responses. At low temporal frequencies, the variance in timing was small, but at higher frequencies, all timings were well represented. The timing data thus matched the temporal frequency tuning of DS. Kittens show DS at high temporal frequencies because of the abundance of inputs with different timing at high frequencies. As cells in the LGN mature, more low-frequency timing differences become available to the cortex, allowing DS at low frequencies to become possible for more cortical cells.
Figures
References
-
- Albrecht DG, Geisler WS. Motion selectivity and the contrast-response function of simple cells in the visual cortex. Vis Neurosci. 1991;7:531–546. - PubMed
-
- Beidenbach MA, Stevens CF. Electrical activity in cat olfactory cortex produced by synchronous orthodromic volleys. J Neurophysiol. 1969;32:193–203. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous