

Plasmodium sporozoite invasion into insect and mammalian cells is directed by the same dual binding system
- PMID: 11927544
- PMCID: PMC125935
- DOI: 10.1093/emboj/21.7.1597
Plasmodium sporozoite invasion into insect and mammalian cells is directed by the same dual binding system
Abstract
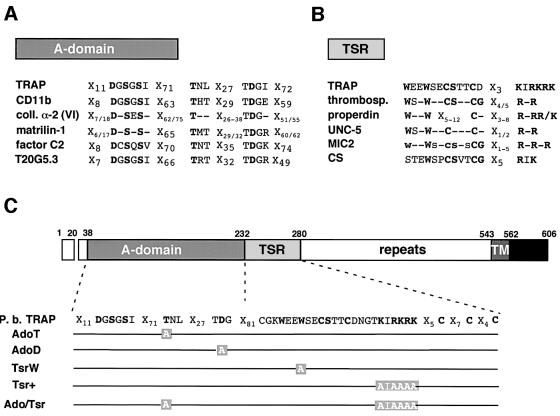
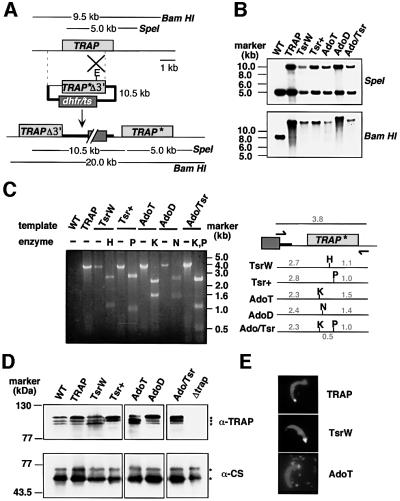
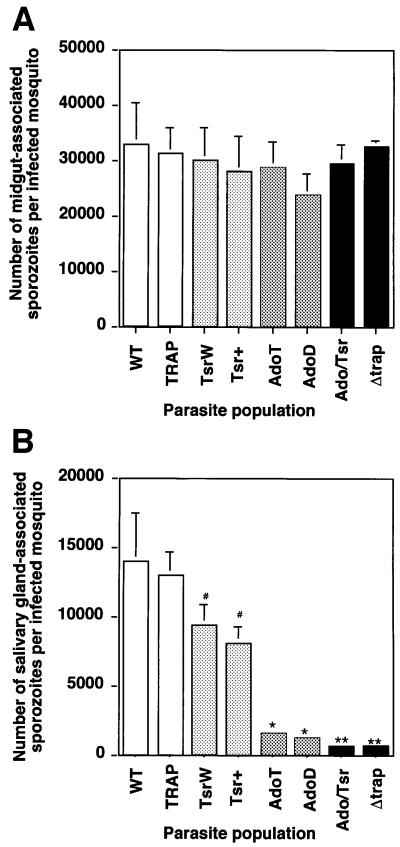
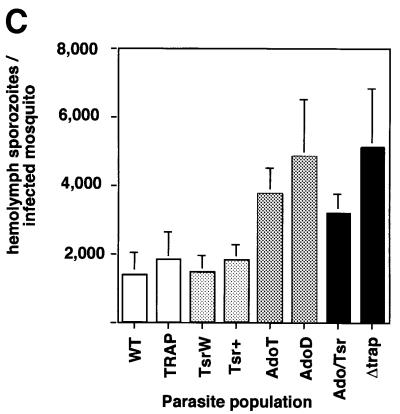
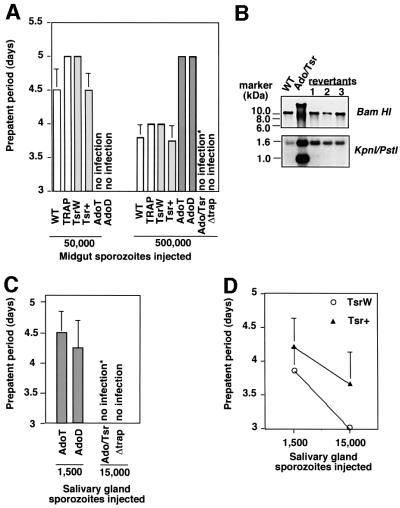
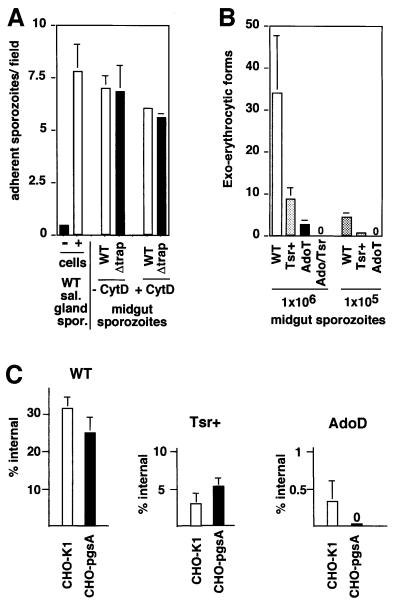
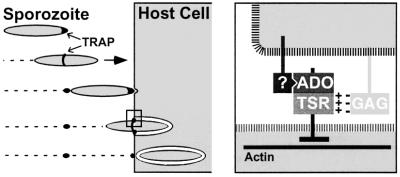
Plasmodium sporozoites, the transmission form of the malaria parasite, successively invade salivary glands in the mosquito vector and the liver in the mammalian host. Sporozoite capacity to invade host cells is mechanistically related to their ability to glide on solid substrates, both activities depending on the transmembrane protein TRAP. Here, we show that loss-of- function mutations in two adhesive modules of the TRAP ectodomain, an integrin-like A-domain and a thrombospondin type I repeat, specifically decrease sporozoite invasion of host cells but do not affect sporozoite gliding and adhesion to cells. Irrespective of the target cell, i.e. in mosquitoes, rodents and cultured human or hamster cells, sporozoites bearing mutations in one module are less invasive, while those bearing mutations in both modules are non-invasive. In Chinese hamster ovary cells, the TRAP modules interact with distinct cell receptors during sporozoite invasion, and thus act as independently active pass keys. As these modules are also present in other members of the TRAP family of proteins in Apicomplexa, they may account for the capacity of these parasites to enter many cell types of phylogenetically distant origins.
Figures
References
-
- Bernfield M., Gotte,M., Park,P.W., Reizes,O., Fitzgerald,M.L., Lincecum,J. and Zako,M. (1999) Functions of cell surface proteoglycans. Annu. Rev. Biochem., 68, 729–777. - PubMed
-
- Carruthers V.B. and Sibley,L.D. (1999) Mobilization of intracellular calcium stimulates microneme discharge in Toxoplasma gondii. Mol. Microbiol., 31, 421–428. - PubMed
-
- Cerami C., Frevert,U., Sinnis,P., Takacs,B., Clavijo,P., Santos,M.J. and Nussenzweig,V. (1992) The basolateral domain of the hepatocyte plasma membrane bears receptors for the circumsporozoite protein of Plasmodium falciparum sporozoites. Cell, 70, 1021–1033. - PubMed
-
- Dobrowolski J.M. and Sibley,L.D. (1996) Toxoplasma invasion of mammalian cells is powered by the actin cytoskeleton of the parasite. Cell, 84, 933–939. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
