

Control of the propagation of dendritic low-threshold Ca(2+) spikes in Purkinje cells from rat cerebellar slice cultures
- PMID: 11927669
- PMCID: PMC2290220
- DOI: 10.1113/jphysiol.2001.013294
Control of the propagation of dendritic low-threshold Ca(2+) spikes in Purkinje cells from rat cerebellar slice cultures
Abstract
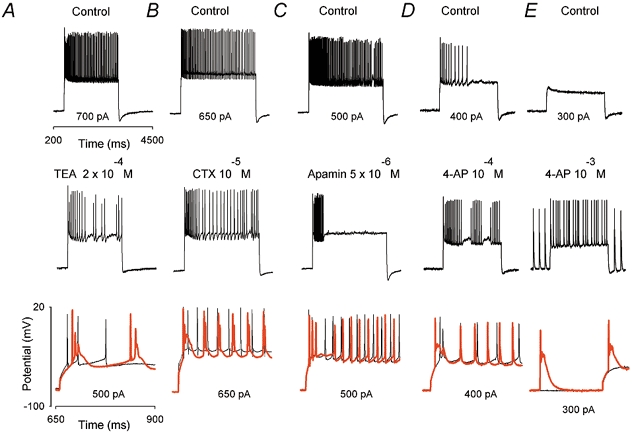
To investigate the ionic mechanisms controlling the dendrosomatic propagation of low-threshold Ca(2+) spikes (LTS) in Purkinje cells (PCs), somatically evoked discharges of action potentials (APs) were recorded under current-clamp conditions. The whole-cell configuration of the patch-clamp method was used in PCs from rat cerebellar slice cultures. Full blockade of the P/Q-type Ca(2+) current revealed slow but transient depolarizations associated with bursts of fast Na(+) APs. These can occur as a single isolated event at the onset of current injection, or repetitively (i.e. a slow complex burst). The initial transient depolarization was identified as an LTS Blockade of P/Q-type Ca(2+) channels increased the likelihood of recording Ca(2+) spikes at the soma by promoting dendrosomatic propagation. Slow rhythmic depolarizations shared several properties with the LTS (kinetics, activation/inactivation, calcium dependency and dendritic origin), suggesting that they correspond to repetitively activated dendritic LTS, which reach the soma when P/Q channels are blocked. Somatic LTS and slow complex burst activity were also induced by K(+) channel blockers such as TEA (2.5 x 10(-4) M) charybdotoxin (CTX, 10(-5) M), rIberiotoxin (10(-7) M), and 4-aminopyridine (4-AP, 10(-3) M), but not by apamin (10(-4) M). In the presence of 4-AP, slow complex burst activity occurred even at hyperpolarized potentials (-80 mV). In conclusion, we suggest that the propagation of dendritic LTS is controlled directly by 4-AP-sensitive K(+) channels, and indirectly modulated by activation of calcium-activated K(+) (BK) channels via P/Q-mediated Ca(2+) entry. The slow complex burst resembles strikingly the complex spike elicited by climbing fibre stimulation, and we therefore propose, as a hypothesis, that dendrosomatic propagation of the LTS could underlie the complex spike.
Figures







References
-
- Cerminara NL, Rawson JA. Distribution of calcium-activated potassium channels in the cerebellum of the rat. Society of Neurosciences Abstracts. 2000;255.6:73.
-
- Coetzee WA, Amarillo Y, Chiu J, Chow A, Lau D, McCormack T, Moreno H, Nadal MS, Ozaita A, Pountney D, Saganich M, Vega-Saenz de Miera E, Rudy B. Molecular diversity of K+ channels. Annals of the New York Academy of Sciences. 1999;29:233–285. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous