

Involvement of the matrix protein in attachment of porcine reproductive and respiratory syndrome virus to a heparinlike receptor on porcine alveolar macrophages
- PMID: 11932397
- PMCID: PMC155060
- DOI: 10.1128/jvi.76.9.4312-4320.2002
Involvement of the matrix protein in attachment of porcine reproductive and respiratory syndrome virus to a heparinlike receptor on porcine alveolar macrophages
Abstract
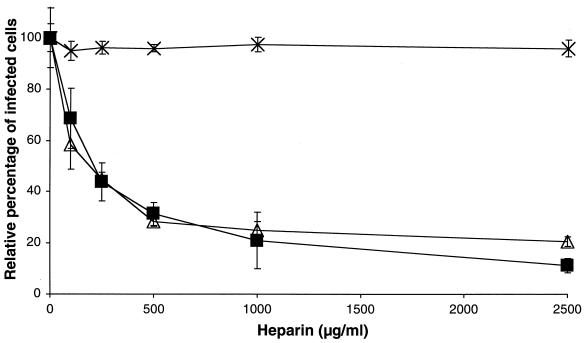
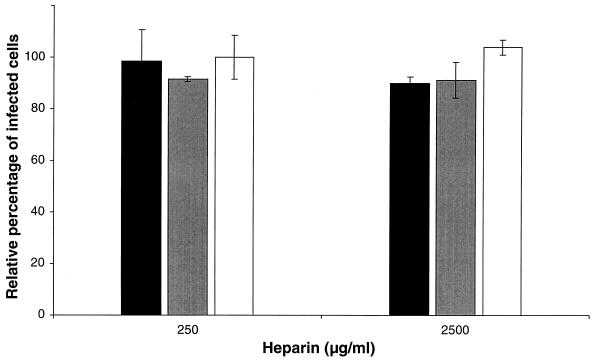
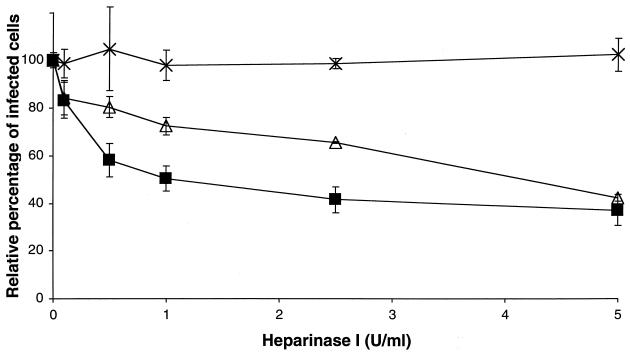
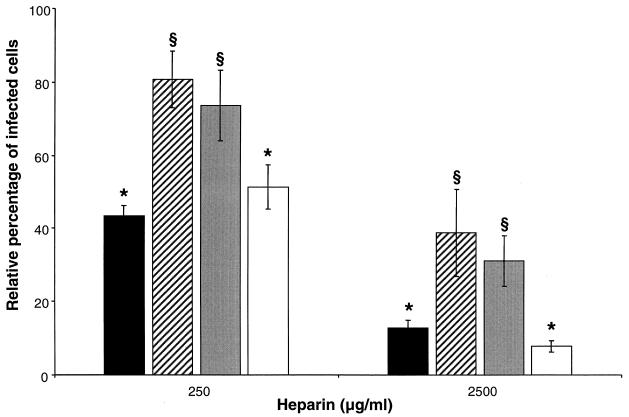
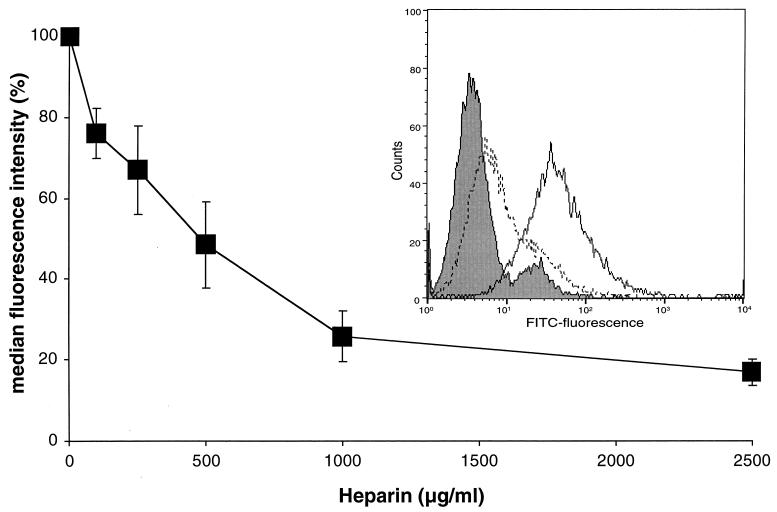
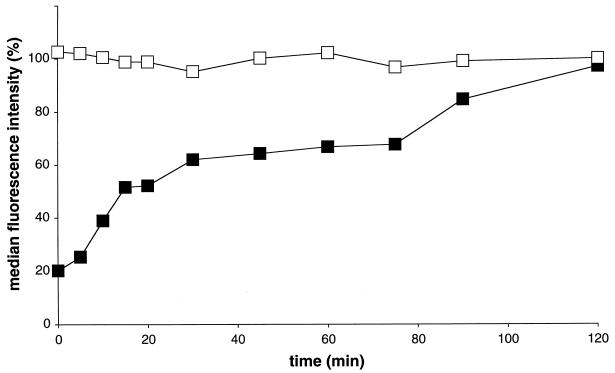
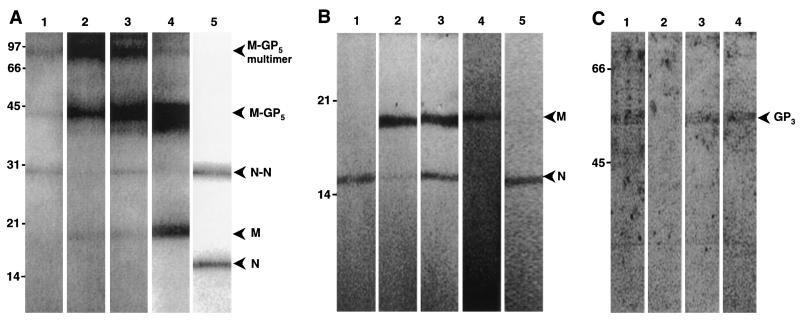
The porcine reproductive and respiratory syndrome virus (PRRSV) has a very restricted tropism for well-differentiated cells of the monocyte-macrophage lineage, which is probably determined by specific receptors on these cells. In this study, the importance of heparinlike molecules on porcine alveolar macrophages (PAM) for PRRSV infection was determined. Heparin interacted with the virus and reduced infection of PAM up to 92 or 88% for the American and European types of PRRSV, respectively. Other glycosaminoglycans, similar to heparin, had no significant effect on infection while heparinase treatment of PAM resulted in a significant reduction of the infection. Analysis of infection kinetics showed that PRRSV attachment to heparan sulfate occurs early in infection. A heparin-sensitive binding step was observed which converted completely into a heparin-resistant binding after 120 min at 4 degrees C. Using heparin-affinity chromatography and sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), it was observed that the structural matrix (M) and nucleocapsid (N) proteins attached to heparin. Nonreducing SDS-PAGE revealed that M bound to heparin mainly as a complex with glycoprotein GP(5) and that the N protein bound to heparin as a homodimer. GP(3), which was identified as a minor structural protein of European types of PRRSV, did not bind to heparin. Since the N protein is not exposed on the virion surface, it was concluded that the structural M protein and the M-GP(5) complex contribute to PRRSV attachment on a heparinlike receptor on PAM. This is the first report that identifies a PRRSV ligand for a cell surface heparinlike receptor on PAM.
Figures
References
-
- Benfield, D. A., E. Nelson, J. E. Collins, L. Harris, S. M. Goyal, D. Robinson, W. T. Christianson, R. B. Morrison, D. Gorcyca, and D. Chladek. 1992. Characterisation of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet. Diagn. Investig. 4:127-133. - PubMed
-
- Bernfield, M., M. Götte, P. W. Park, O. Reizes, M. L. Fitzgerald, J. Lincecum, and M. Zako. 1999. Functions of cell surface heparan sulphate proteoglycans. Annu. Rev. Biochem. 68:729-777. - PubMed
-
- Cavanagh, D. 1997. Nidovirales: a new order comprising Coronaviridae and Arteriviridae. Arch. Virol. 142:629-633. - PubMed
-
- Collins, J. E., D. A. Benfield, W. T. Christianson, L. Harris, J. C. Hennings, D. P. Shaw, S. M. Goyal, S. McCullough, R. B. Morrison, H. S. Joo, D. Gorcyca, and D. Chladek. 1992. Isolation of swine infertility and respiratory syndrome virus (isolate ATCC VR-2332) in North America and experimental reproduction of the disease in gnotobiotic pigs. J. Vet. Diagn. Investig. 4:117-126. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
