

Role of DIVARICATA in the control of dorsoventral asymmetry in Antirrhinum flowers
- PMID: 11937495
- PMCID: PMC186332
- DOI: 10.1101/gad.221002
Role of DIVARICATA in the control of dorsoventral asymmetry in Antirrhinum flowers
Abstract
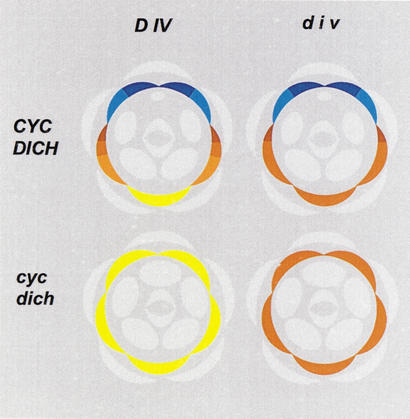
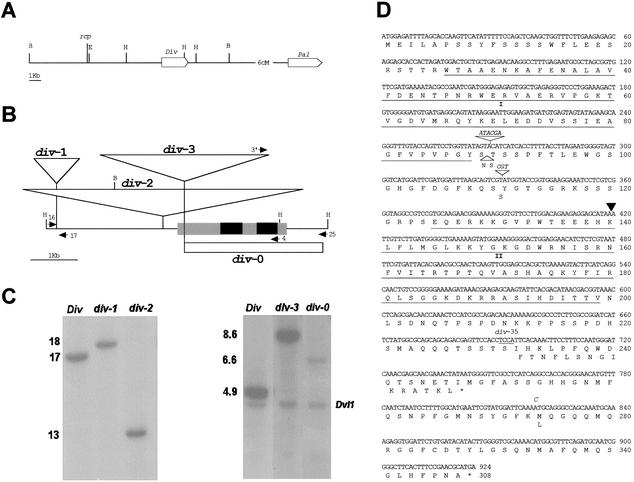
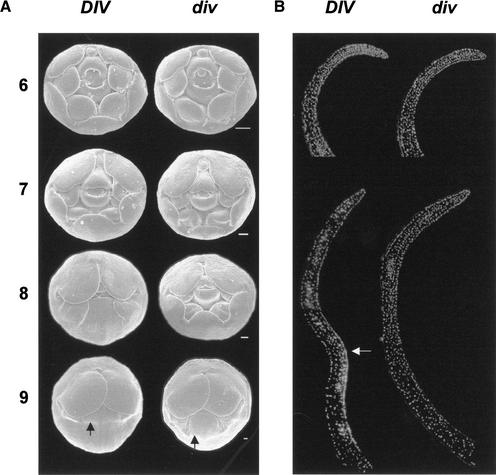
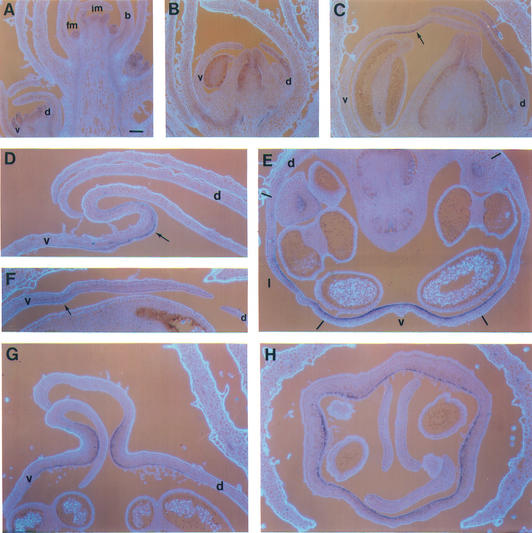
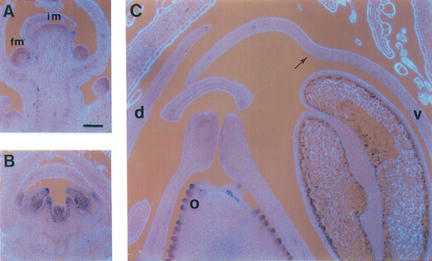
Dorsoventral asymmetry of the Antirrhinum corolla depends on expression of the CYC and DICH genes in dorsal petals. One role of these genes is to inhibit DIVARICATA (DIV), a determinant of ventral identity. Therefore, in cyc;dich double mutants ventral identity spreads all around the flower. We show that DIV encodes a protein belonging to the MYB family of transcription factors. Early on in corolla development, DIV affects specifically the growth of ventral and lateral petals but is transcribed in all petals. Analysis of a closely related gene suggests that the lack of effect on dorsal petals is not due to redundancy. More likely, therefore, DIV is regulated posttranscriptionally through a mechanism that depends on CYC and DICH. Later on, DIV affects growth and cell types and is transcribed mostly in a single layer of cells of ventral and lateral petals. This late pattern may itself depend on DIV activity because it fails to be established in a transcribed but inactive div mutant and, conversely, spreads all around the flower in cyc;dich double mutants.
Figures


References
-
- Almeida J, Carpenter R, Robbins TP, Martin C, Coen ES. Genetic interactions underlying flower color patterns in Antirrhinum majus. Genes & Dev. 1989;3:1758–1767. - PubMed
-
- Almeida J, Rocheta M, Galego L. Genetic control of flower shape in Antirrhinum majus. Development. 1997;124:1387–1392. - PubMed
-
- Altschul SF, Gish W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases