

Enhanced expression of the transcription factor Nrf2 by cancer chemopreventive agents: role of antioxidant response element-like sequences in the nrf2 promoter
- PMID: 11940647
- PMCID: PMC133753
- DOI: 10.1128/MCB.22.9.2883-2892.2002
Enhanced expression of the transcription factor Nrf2 by cancer chemopreventive agents: role of antioxidant response element-like sequences in the nrf2 promoter
Abstract
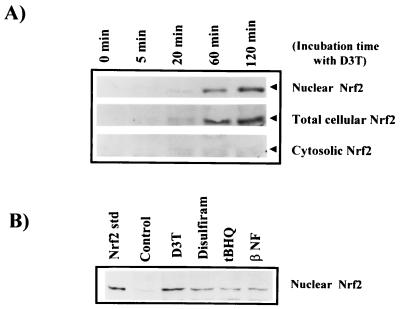
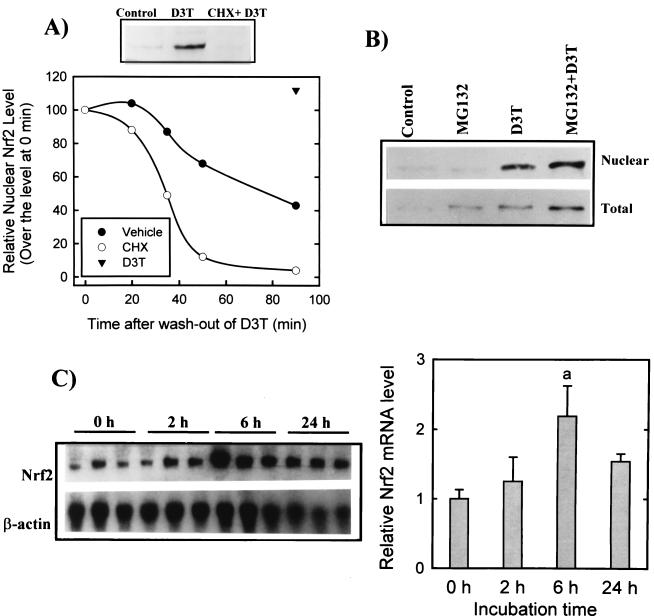
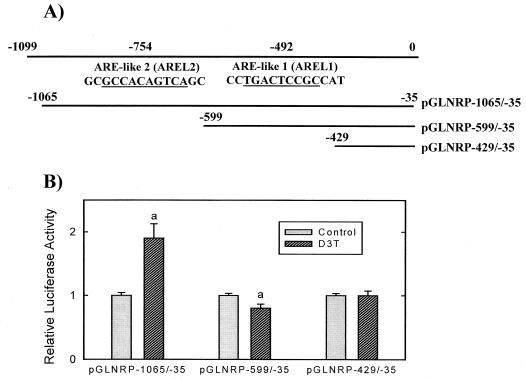
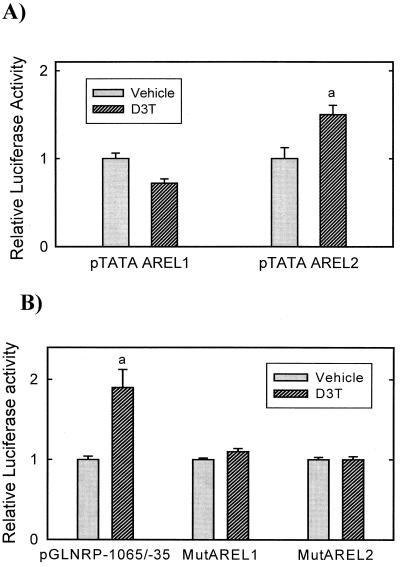
Induction of phase 2 enzymes, which neutralize reactive electrophiles and act as indirect antioxidants, is an important mechanism for protection against carcinogenesis. The transcription factor Nrf2, which binds to the antioxidant response element (ARE) found in the upstream regulatory region of many phase 2 genes, is essential for the induction of these enzymes. We have investigated the effect of the potent enzyme inducer and anticarcinogen 3H-1,2-dithiole-3-thione (D3T) on the fate of Nrf2 in murine keratinocytes. Both total and nuclear Nrf2 levels increased rapidly and persistently after treatment with D3T but could be blocked by cotreatment with cycloheximide. Nrf2 mRNA levels increased approximately 2-fold 6 h after D3T treatment. To examine the transcriptional activation of Nrf2 by D3T, the proximal region (1 kb) of the nrf2 promoter was isolated. Deletion and mutagenesis analyses demonstrated that nrf2 promoter-luciferase reporter activity was enhanced by treatment with D3T and that ARE-like sequences were required for this activation. Gel shift assays with nuclear extracts from PE cells indicated that common factors bind to typical AREs and the ARE-like sequences of the nrf2 promoter. Direct binding of Nrf2 to its own promoter was demonstrated by chromatin immunoprecipitation assay. Overexpression of Nrf2 increased the activity of the nrf2 promoter-luciferase reporter, while expression of mutant Nrf2 protein repressed activity. Thus, Nrf2 appears to autoregulate its own expression through an ARE-like element located in the proximal region of its promoter, leading to persistent nuclear accumulation of Nrf2 and protracted induction of phase 2 genes in response to chemopreventive agents.
Figures
References
-
- Alam, J., D. Stewart, C. Touchard, S. Boinapally, A. M. K. Choi, and J. L. Cook. 1999. Nrf2, a cap'n'collar transcription factor, regulates induction of the heme oxygenase-1 gene. J. Biol. Chem. 274:26071-26078. - PubMed
-
- Angel, P., K. Hattori, T. Smeal, and M. Karin. 1988. The jun proto-oncogene is positively autoregulated by its product, Jun/AP-1. Cell 55:875-885. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases