

Altered p27(Kip1) phosphorylation, localization, and function in human epithelial cells resistant to transforming growth factor beta-mediated G(1) arrest
- PMID: 11940657
- PMCID: PMC133745
- DOI: 10.1128/MCB.22.9.2993-3002.2002
Altered p27(Kip1) phosphorylation, localization, and function in human epithelial cells resistant to transforming growth factor beta-mediated G(1) arrest
Abstract
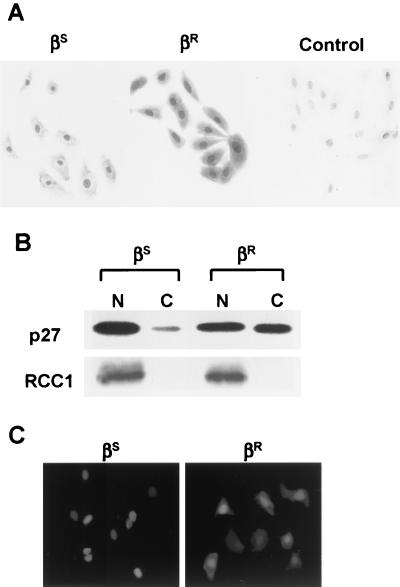
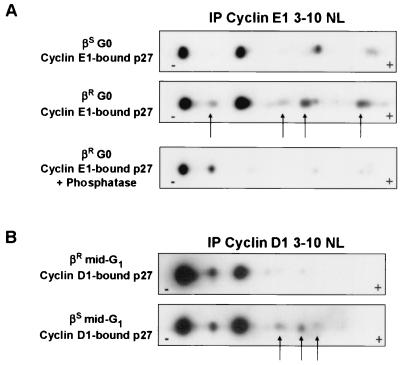
p27(Kip1) is an important effector of G(1) arrest by transforming growth factor beta (TGF-beta). Investigations in a human mammary epithelial cell (HMEC) model, including cells that are sensitive (184(S)) and resistant (184A1L5(R)) to G(1) arrest by TGF-beta, revealed aberrant p27 regulation in the resistant cells. Cyclin E1-cyclin-dependent kinase 2 (cdk2) and cyclin A-cdk2 activities were increased, and p27-associated kinase activity was detected in 184A1L5(R) cells. p27 from 184A1L5(R) cells was localized to both nucleus and cytoplasm, showed an altered profile of phosphoisoforms, and had a reduced ability to bind and inhibit cyclin E1-cdk2 in vitro when compared to p27 from the sensitive 184(S) cells. In proliferating 184A1L5(R) cells, more p27 was associated with cyclin D1-cdk4 complexes than in 184(S). While TGF-beta inhibited the formation of cyclin D1-cdk4-p27 complexes in 184(S) cells, it did not inhibit the assembly of cyclin D1-cdk4-p27 complexes in the resistant 184A1L5(R) cells. p27 phosphorylation changed during cell cycle progression, with cyclin E1-bound p27 in G(0) showing a different phosphorylation pattern from that of cyclin D1-bound p27 in mid-G(1). These data suggest a model in which TGF-beta modulates p27 phosphorylation from its cyclin D1-bound assembly phosphoform to an alternate form that binds tightly to inhibit cyclin E1-cdk2. Altered phosphorylation of p27 in the resistant 184A1L5(R) cells may favor the binding of p27 to cyclin D1-cdk4 and prevent its accumulation in cyclin E1-cdk2 in response to TGF-beta.
Figures





References
-
- Carrano, A. C., E. Eytan, A. Hershko, and M. Pagano. 1999. SKP2 is required for ubiquitin-mediated degradation of the CDK inhibitor p27. Nat. Cell Biol. 1:193-199. - PubMed
-
- Catzavelos, C., N. Bhattacharya, Y. C. Ung, J. A. Wilson, L. Roncari, C. Sandhu, P. Shaw, H. Yeger, I. Morava-Protzner, L. Kapusta, E. Franssen, K. I. Pritchard, and J. M. Slingerland. 1997. Decreased levels of the cell-cycle inhibitor p27Kip1 protein: prognostic implications in primary breast cancer. Nat. Med. 3:227-230. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous