

Initiation sites are distributed at frequent intervals in the Chinese hamster dihydrofolate reductase origin of replication but are used with very different efficiencies
- PMID: 11940663
- PMCID: PMC133756
- DOI: 10.1128/MCB.22.9.3053-3065.2002
Initiation sites are distributed at frequent intervals in the Chinese hamster dihydrofolate reductase origin of replication but are used with very different efficiencies
Abstract
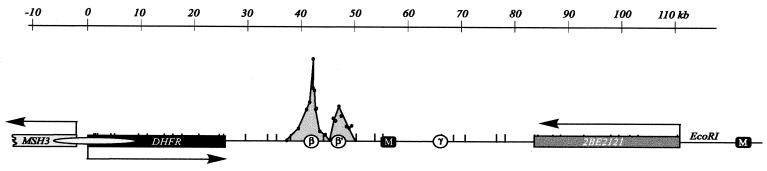
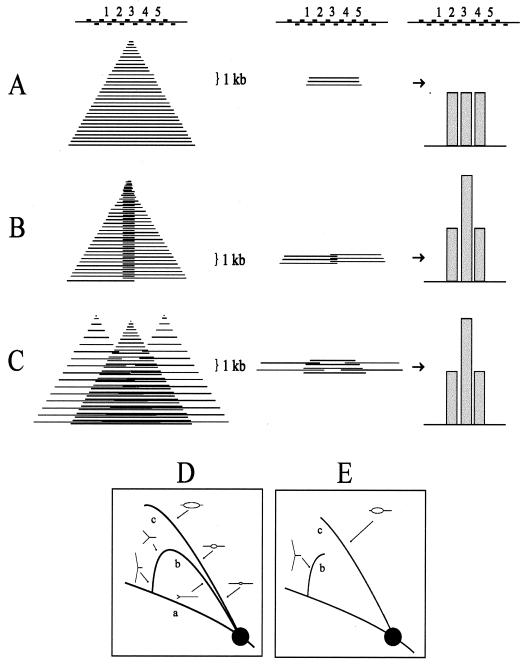
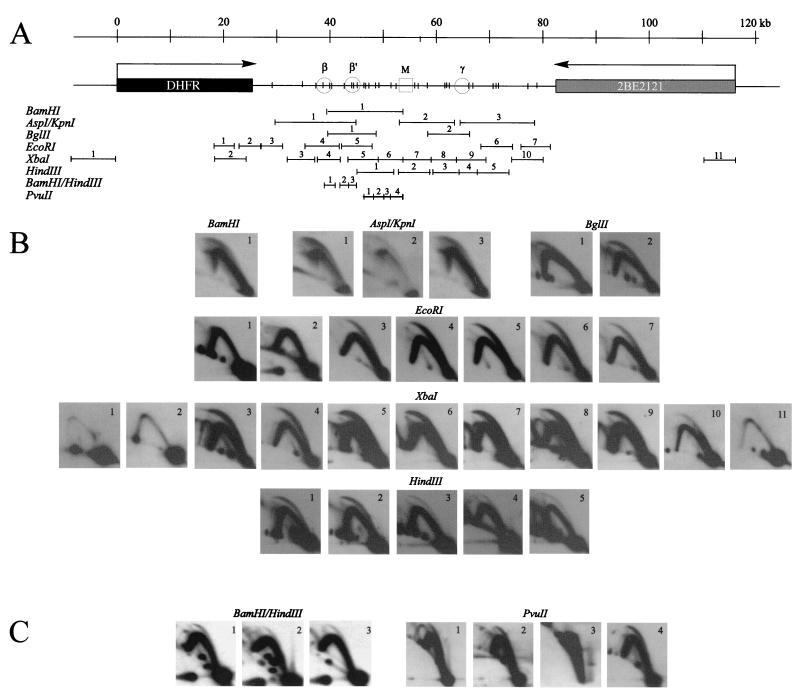
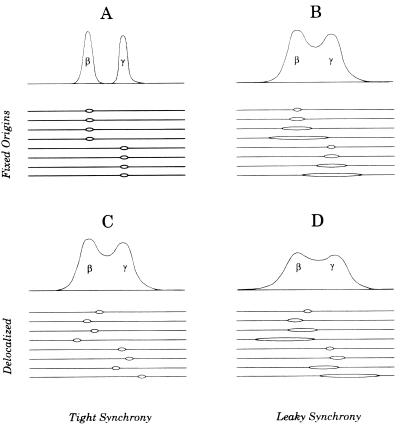
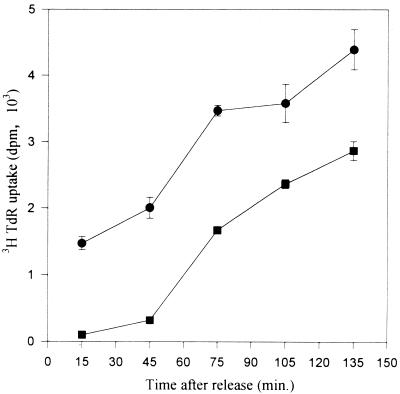
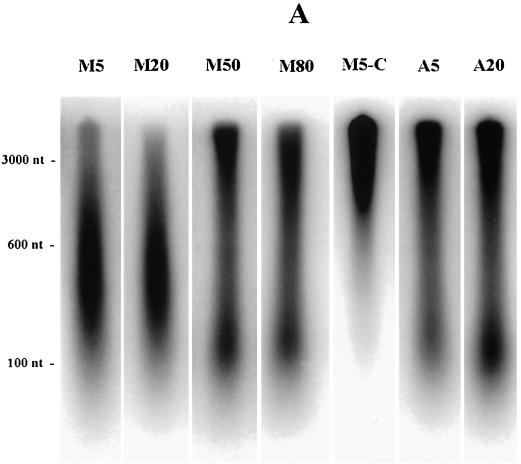
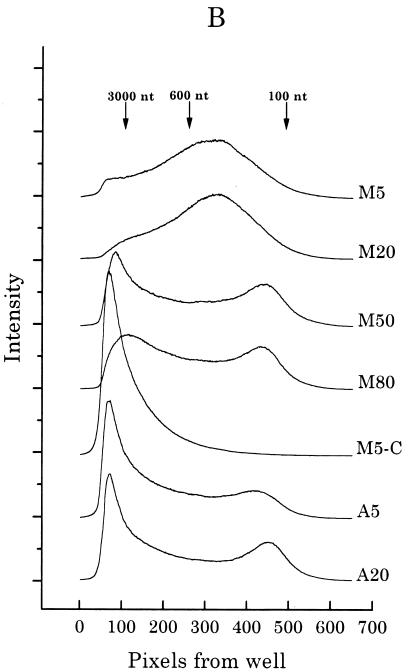
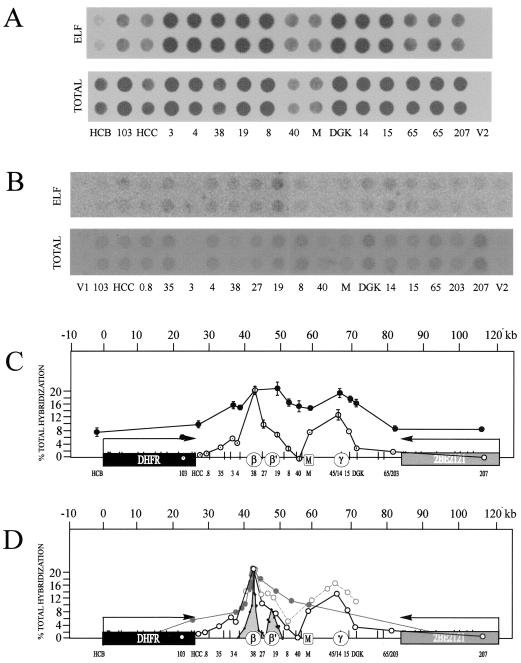
Previous radiolabeling and two-dimensional (2-D) gel studies of the dihydrofolate reductase (DHFR) domain of Chinese hamster cells have suggested that replication can initiate at any one of a very large number of inefficient sites scattered throughout the 55-kb intergenic spacer region, with two broad subregions (ori-beta and ori-gamma) preferred. However, high-resolution analysis by a PCR-based nascent strand abundance assay of the 12-kb subregion encompassing ori-beta has suggested the presence of a relatively small number of fixed, highly efficient initiation sites distributed at infrequent intervals that correspond to genetic replicators. To attempt to reconcile these observations, two different approaches were taken in the present study. In the first, neutral-neutral 2-D gel analysis was used to examine replication intermediates in 31 adjacent and overlapping restriction fragments in the spacer, ranging in size from 1.0 to 18 kb. Thirty of 31 fragments displayed the complete bubble arcs characteristic of centered origins. Taking into account overlapping fragments, these data suggest a minimum of 14 individual start sites in the spacer. In the second approach, a quantitative early labeled fragment hybridization assay was performed in which radioactive origin-containing DNA 300 to 1,000 nucleotides in length was synthesized in the first few minutes of the S period and used to probe 15 clones distributed throughout the intergenic spacer but separated on average by more than 1,000 bp. This small nascent DNA fraction hybridized to 14 of the 15 clones, ranging from just above background to a maximum at the ori-beta locus. The only silent region detected was a small fragment lying just upstream from a centered matrix attachment region--the same region that was also negative for initiation by 2-D gel analysis. Results of both approaches suggest a minimum of approximately 20 initiation sites in the spacer (two of them being ori-beta and ori-gamma), with ori-beta accounting for a maximum of approximately 20% of initiations occurring in the spacer. We believe that the results of all experimental approaches applied to this locus so far can be fitted to a model in which the DHFR origin consists of a 55-kb intergenic zone of potential sites that are used with very different efficiencies and which are separated in many cases by a few kilobases or less.
Figures
References
-
- Abdurashidova, G., M. Deganuto, R. Klima, S. Riva, G. Biamonti, M. Giacca, and A. Falaschi. 2000. Start sites of bidirectional DNA synthesis at the human lamin B2 origin. Science 287:2023-2026. - PubMed
-
- Bielinsky, A. K., and S. A. Gerbi. 1998. Discrete start sites for DNA synthesis in the yeast ARS1 origin. Science 279:95-98. - PubMed
-
- Brewer, B. J., and W. L. Fangman. 1987. The localization of replication origins on ARS plasmids in S. cerevisiae. Cell 51:463-471. - PubMed
-
- Burhans, W. C., L. T. Vassilev, M. S. Caddle, N. H. Heintz, and M. L. DePamphilis. 1990. Identification of an origin of bidirectional DNA replication in mammalian chromosomes. Cell 62:955-965. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources