

Activation of mislocalized opsin kills rod cells: a novel mechanism for rod cell death in retinal disease
- PMID: 11943854
- PMCID: PMC122826
- DOI: 10.1073/pnas.072557799
Activation of mislocalized opsin kills rod cells: a novel mechanism for rod cell death in retinal disease
Abstract
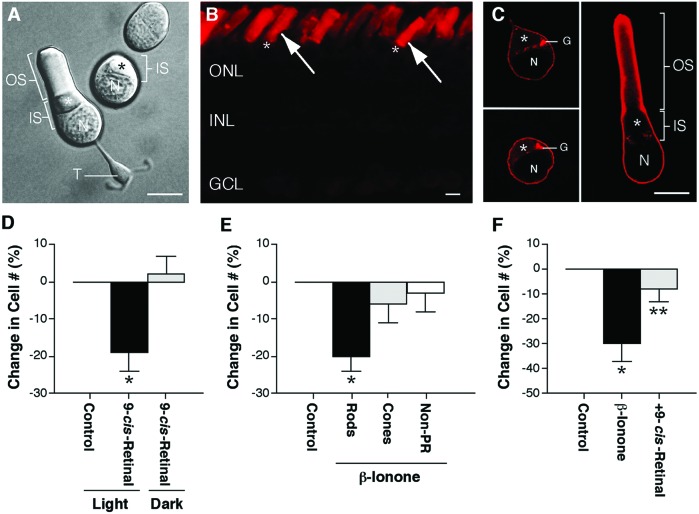
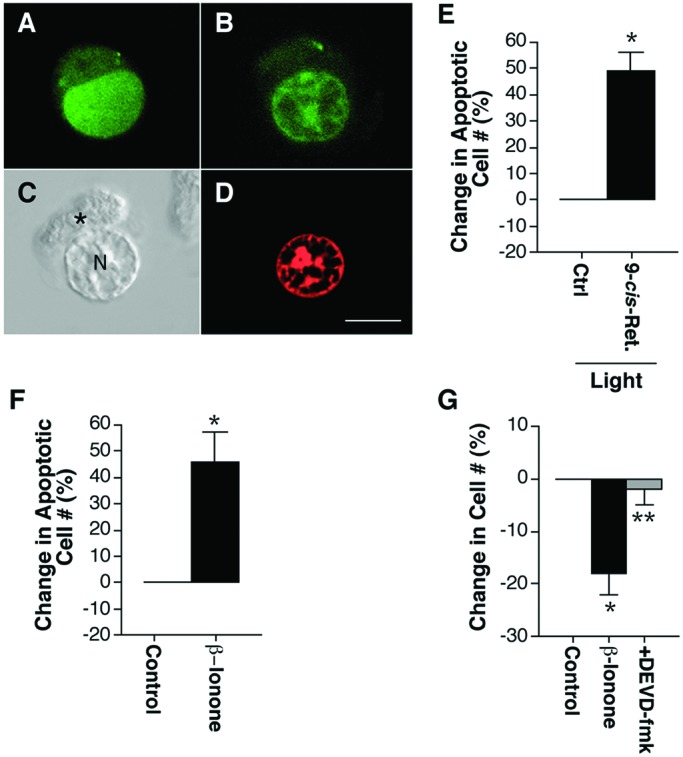
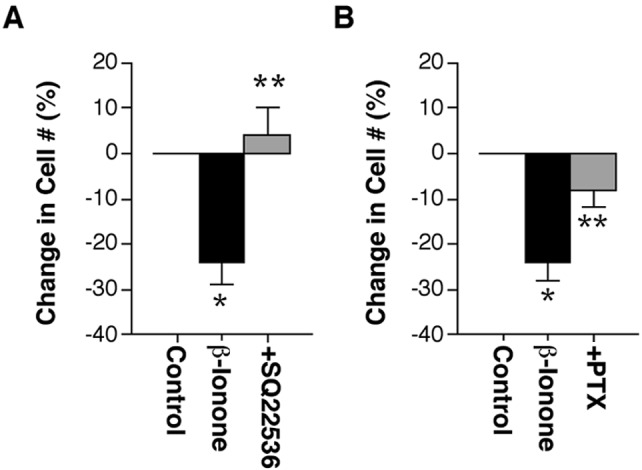
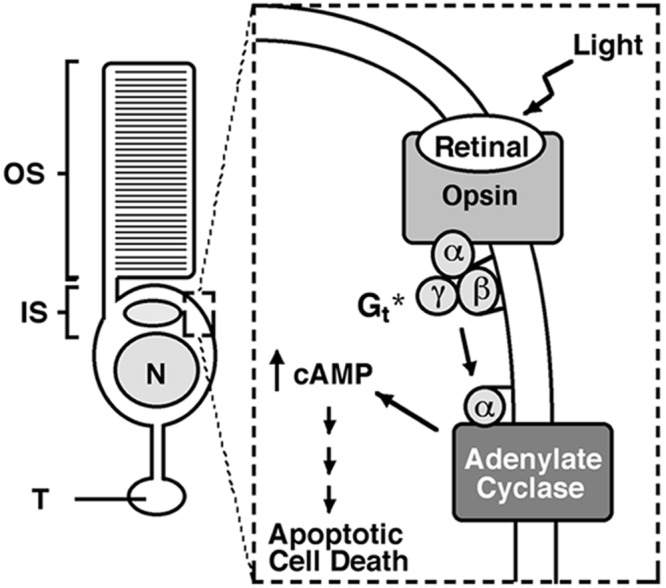
Rod photoreceptors are highly compartmentalized sensory neurons that maintain strict ultrastructural and molecular polarity. Structural subdivisions include the outer segment, inner segment, cell body, and synaptic terminal. The visual pigment rhodopsin is found predominantly in membranes of the rod cell outer segment but becomes mislocalized, appearing throughout the plasma membrane of the cell in many retinal diseases and injuries. Currently, there is no known link between rhodopsin redistribution and rod cell death. We propose that activation of mislocalized rhodopsin kills rod cells by stimulating normally inaccessible signaling pathways. This hypothesis was tested in primary retinal cell cultures, which contain photoreceptors. In rod photoreceptors, opsin immunofluorescence occurred throughout the rod cell plasma membrane. Activation of this mislocalized opsin by photostimulation after formation of isorhodopsin or by incubation with beta-ionone (opsin agonist) killed 19-30% of rod cells. Rod cell death was apoptotic, as indicated by marked chromatin condensation and the requirement for caspase-3 activation. Rod cell death could be induced by forskolin (adenylate cyclase agonist), and conversely, beta-ionone-induced cell death could be blocked by cotreatment with SQ22536 (an adenylate cyclase inhibitor). Pertussis toxin (a G protein inhibitor) also blocked beta-ionone-induced cell death. The data support a mechanism by which activation of mislocalized opsin initiates apoptotic rod cell death through G protein stimulation of adenylate cyclase.
Figures
References
-
- Schmidt S Y. In: Handbook of Neurochemistry, Volume 10. Lajthe A, editor. New York: Plenum; 1985. pp. 461–507.
-
- Li Z-Y, Milam A H. In: Degenerative Diseases of the Retina. Anderson R E, editor. New York: Plenum; 1995. pp. 1–8.
-
- Cook B, Lewis G P, Fisher S K, Adler R. Invest Ophthalmol Visual Sci. 1995;36:990–996. - PubMed
-
- Abler A S, Chang C J, Ful J, Tso M O M, Lam T T. Res Commun Mol Pathol Pharmacol. 1996;92:177–189. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
