

Grx5 is a mitochondrial glutaredoxin required for the activity of iron/sulfur enzymes
- PMID: 11950925
- PMCID: PMC102255
- DOI: 10.1091/mbc.01-10-0517
Grx5 is a mitochondrial glutaredoxin required for the activity of iron/sulfur enzymes
Abstract
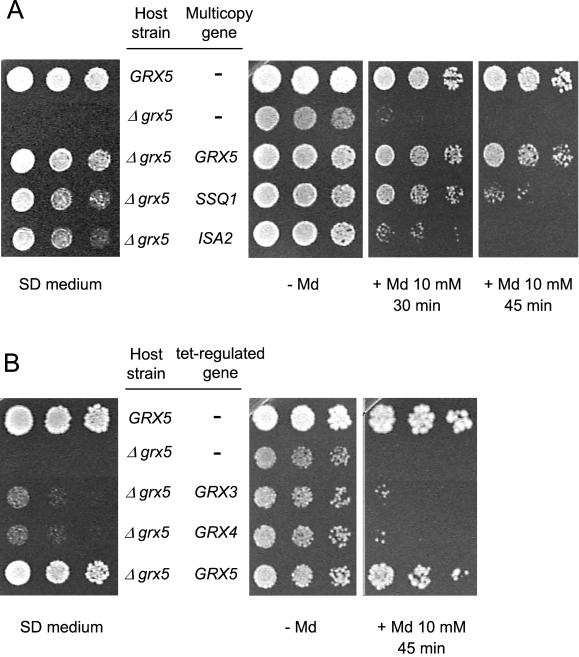
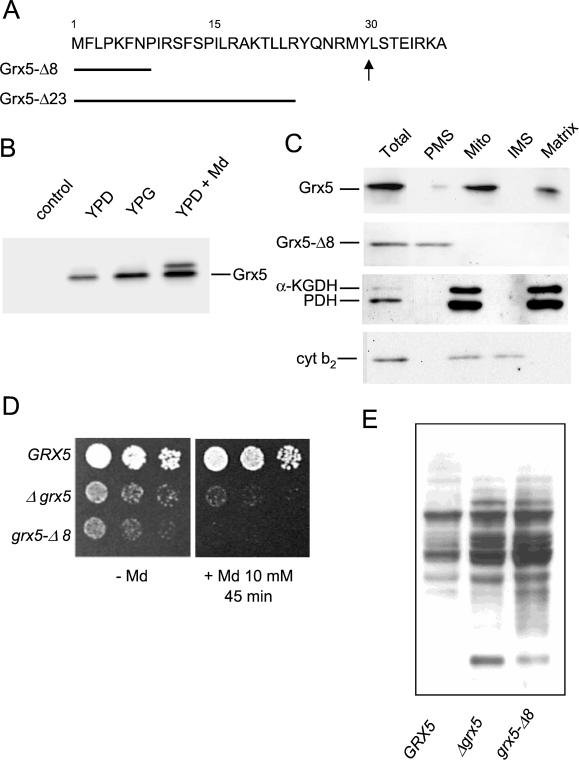
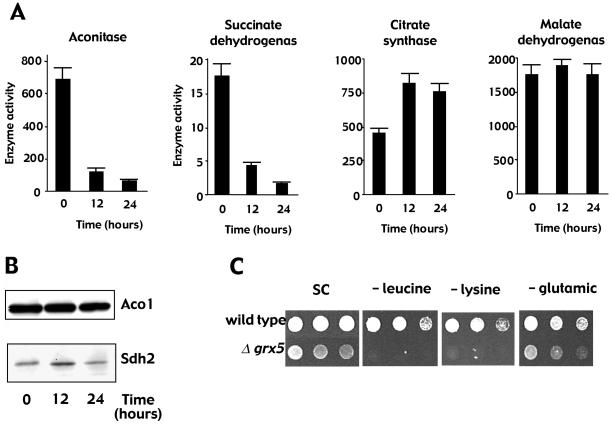
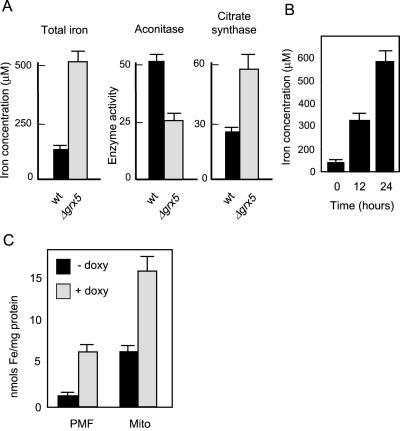
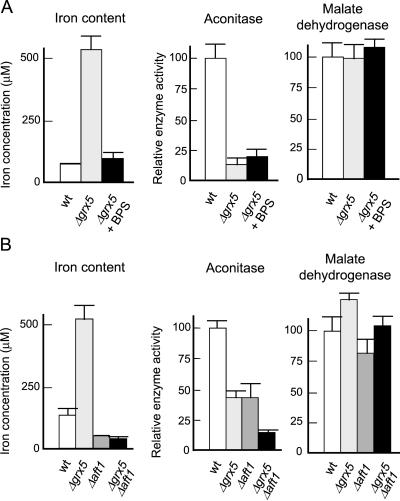
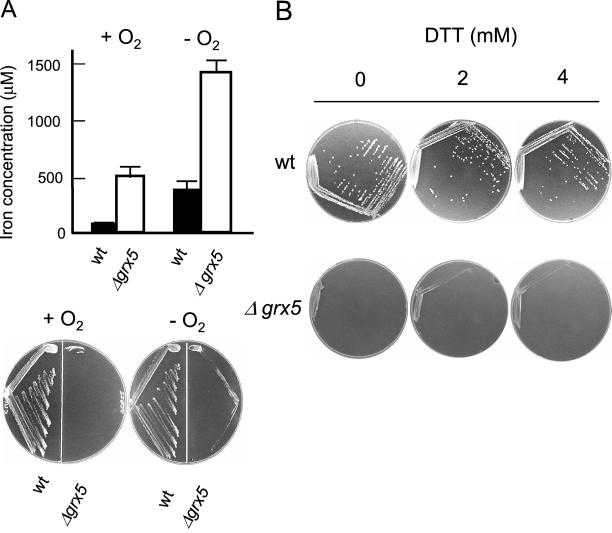
Yeast cells contain a family of three monothiol glutaredoxins: Grx3, 4, and 5. Absence of Grx5 leads to constitutive oxidative damage, exacerbating that caused by external oxidants. Phenotypic defects associated with the absence of Grx5 are suppressed by overexpression of SSQ1 and ISA2, two genes involved in the synthesis and assembly of iron/sulfur clusters into proteins. Grx5 localizes at the mitochondrial matrix, like other proteins involved in the synthesis of these clusters, and the mature form lacks the first 29 amino acids of the translation product. Absence of Grx5 causes: 1) iron accumulation in the cell, which in turn could promote oxidative damage, and 2) inactivation of enzymes requiring iron/sulfur clusters for their activity. Reduction of iron levels in grx5 null mutants does not restore the activity of iron/sulfur enzymes, and cell growth defects are not suppressed in anaerobiosis or in the presence of disulfide reductants. Hence, Grx5 forms part of the mitochondrial machinery involved in the synthesis and assembly of iron/sulfur centers.
Figures
References
-
- Askwith C, Kaplan J. Iron and copper transport in yeast and its relevance to human disease. Trends Biochem Sci. 1998;23:135–138. - PubMed
-
- Babcock M, De Silva D, Oaks R, Davis-Kaplan S, Jiralerspong S, Montermini L, Pandolfo M, Kaplan J. Regulation of mitochondrial iron accumulation by Yfh1p, a putative homolog of frataxin. Science. 1997;276:1709–1712. - PubMed
-
- Barros MH, Nobrega FG. YAH1 of Saccharomyces cerevisiae: a new essential gene that codes for a protein homologous to human adrenodoxin. Gene. 1999;233:197–203. - PubMed
-
- Beinert H, Holm RH, Münck E. Iron-sulfur clusters: nature's modular, multipurpose structures. Science. 1997;277:653–659. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
