

doi: 10.1091/mbc.01-10-0482.
Mutations in Saccharomyces cerevisiae gene SIR2 can have differential effects on in vivo silencing phenotypes and in vitro histone deacetylation activity
Affiliations
- PMID: 11950950
- PMCID: PMC102280
- DOI: 10.1091/mbc.01-10-0482
Item in Clipboard
Mutations in Saccharomyces cerevisiae gene SIR2 can have differential effects on in vivo silencing phenotypes and in vitro histone deacetylation activity
Mol Biol Cell.
2002 Apr.
Abstract
The yeast SIR2 gene and many of its homologs have been identified as NAD(+)-dependent histone deacetylases. To get a broader view of the relationship between the histone deacetylase activity of Sir2p and its in vivo functions we have mutated eight highly conserved residues in the core domain of SIR2. These mutations have a range of effects on the ability of Sir2p to deacetylate histones in vitro and to silence genes at the telomeres and HM loci. Interestingly, there is not a direct correlation between the in vitro and in vivo effects in some of these mutations. We also show that the histone deacetylase activity of Sir2p is necessary for the proper localiztion of the SIR complex to the telomeres.
Figures

Mutagenesis of the core domain of SIR2. (A) Eight amino acid residues that are conserved between bacterial CobB, Saccharomyces SIR2, and mouse SIR2α were mutated to alanine in ySIR2. They correspond to the following residues: Thr-261, Gly-270, Ile-271, Phe-274, Arg-275, Asn-345, Asp-347, and His-364. (B) rSir2p from E. coli was purified over a Ni+ column. The purified protein was run on a polyacrylamide gel to test for purity and stability.
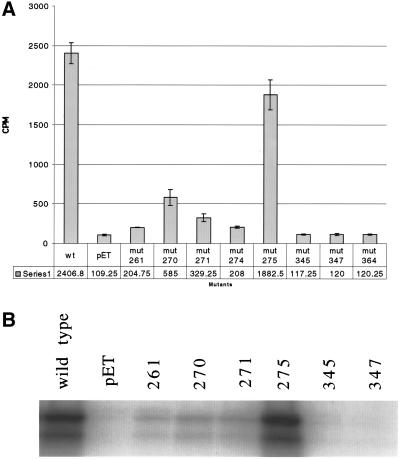
Analysis of Sir2p's enzymatic activities. (A) Tritiated H4 peptide was used to measure the efficiency of the mutants in an NAD+-dependent deacetylation reaction. The peptide was incubated with 1 mM NAD+ and 2 μg of recombinant protein. The graph measures deacetylation activity by counting the amount of tritiated acetate released from the peptide. (B) Capability of the mutant rSir2 proteins to ribosylate histone H3 was measured by incubating recombinant enzyme with [32P]NAD+ and calf histone H3. The products were run out on a polyacrylamide gel and the gel was exposed to film to see whether the histones incorporated label from the [32P]NAD+.
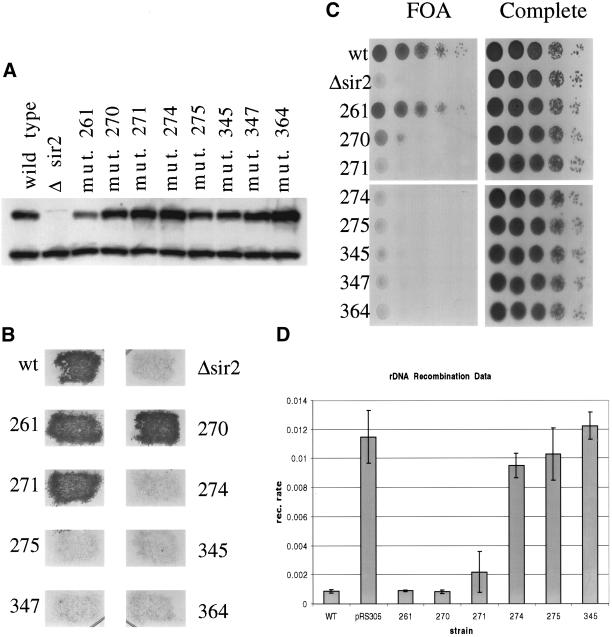
Testing sir2 mutant phenotypes in vivo. (A) Whole cell protein extracts were made from each of the indicated strains. Whole cell extract (15 μg) was run on a polyacrylamide gel and blotted to a polyvinylidene difluoride membrane for Western blot analysis. The blot was probed with anti-Sir2p to measure the level of Sir2p in each of the mutant strains. The upper band is the Sir2p band. (B) To investigate silencing at the HMLα locus, the mutant strains were mated with a mating tester strain and grown on selective media to select for diploids. (C) To test for telomere silencing, SIR2 was mutated in a strain background with the URA3 marker at the end of telomere VII. Each mutant strain was tested for its ability to silence the marker by growing on media containing 5-FOA (a substrate that is toxic to yeast expressing URA3). (D) rDNA recombination rates were measured in a strain background with the ADE2 marker located in the rDNA array. The rate of marker loss was measured for each mutant by counting the number of colonies that lost the marker in the first generation after plating.
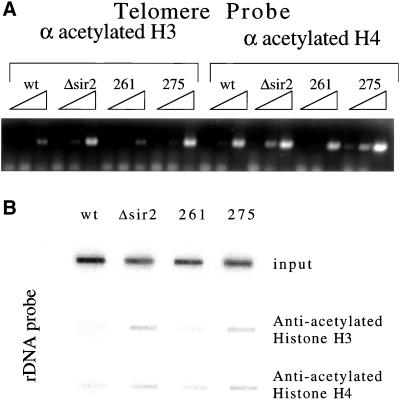
Chromatin IP by using anti-acetylated histone antibody to measure histone acetylation at silent loci. (A) Chromatin IP was used to investigate the acetylation state of the histones at the telomeres in mutants 261 and 275. Threefold dilutions of anti-acetylated H3 and anti-acetylated H4 IP DNA were probed for telomeres by using PCR primers corresponding to the telomere on the right arm of chromosome VI. (B) IP DNA described above was blotted to a membrane by using a slot blot apparatus. The DNA was then probed for rDNA sequences to test the level of acetylation of histone H3 and H4 at the rDNA in the mutant strains.
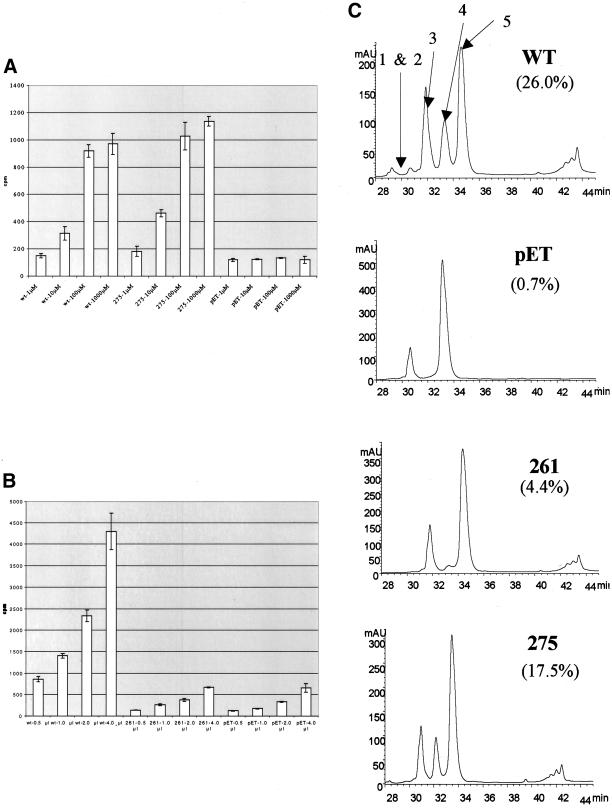
Histone deacetylation activity of mutants under different NAD+ and peptide concentrations. (A) Histone deacetylation activity of mutant 275 along with wild-type Sir2p and no enzyme control was measured using tritiated H4 peptide that was incubated with enzyme and concentrations of NAD+ ranging from 1 μM to 1 mM. Deacetylation activity was measured by counting the amount of free acetate removed during the reaction. (B) Histone deacetylation activity of mutant 261 along with wild-type Sir2p and no enzyme was measured while the amount of tritiated peptide was varied from 0.5 to 4 μl. (C) HPLC chromatograms showing absorbance at 220 nm of products of deacetylation assays with yeast Sir2p, pET (no recombinant protein), mutant 261, and mutant 275 after a 1-h reaction. The efficiencies of the reactions are calculated as a fraction of the areas under peaks 1, 2, and 4 compared with the area under all of the peaks.

Life span of mutant sir2 strains. (A) In an HMLα-deleted background, the life spans of mutant 261 and 270 are the same as wild type, whereas mutant 275 is comparable to Δsir2. The mean life spans are as follows: wild type, 20.7; Δsir2, 11.9; mutant 261, 20.8; mutant 270, 21.4; and mutant 275, 11.9. (B) In an HML + background, the life spans of mutants 261 and 270 are intermediate. The mean life spans are as follows: wild type, 24.5; Δsir2, 11.3; mutant 261, 19.9; and mutant 270, 18.9.
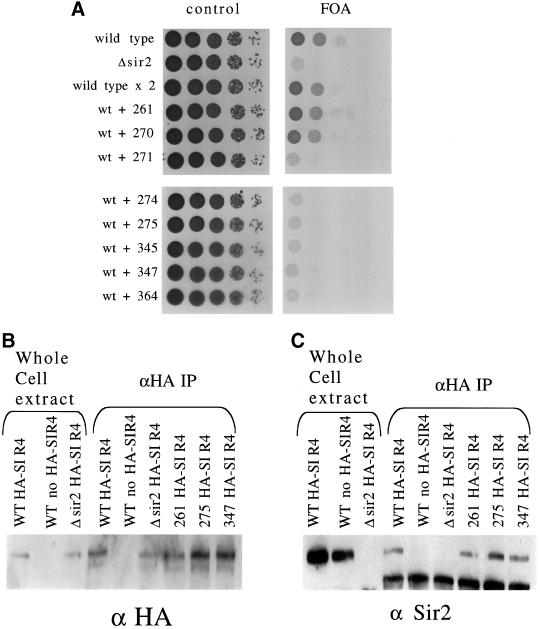
Dominance of mutations and coimmunoprecipitation of SIR2 and SIR4. (A) W303-ARUT strains with a wild-type and mutant copy of SIR2 were spotted onto control synthetic media and 5-FOA plates to test whether the mutations had a dominant effect on telomere silencing. (B) Whole cell protein extract from selected mutant strains (wild type, Δsir2, mutant 261, mutant 275, and mutant 347) with an HA-tagged SIR4 gene was immunoprecipitated with antibodies against the HA epitope. The IP extract along with whole cell extract was run on a polyacrylamide gel and blotted to a polyvinylidene difluoride membrane so that Western blotting analysis could be performed. The membrane was probed using antibody to the HA epitope to measure the efficiency of the pull down. (C) Polyvinylidene difluoride membrane is reprobed with antibody recognizing Sir2p to measure the ability of the mutant Sir2ps to interact with Sir4p in vivo.
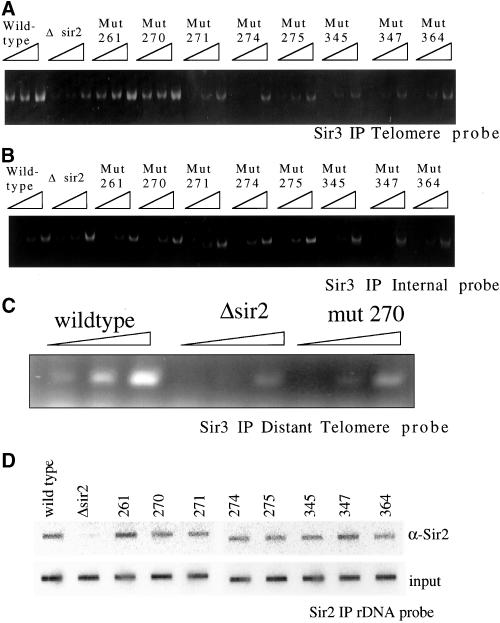
Chromatin IP of Sir complex. Chromatin immunoprecipitation was used to investigate the ability of the Sir complex to localize to the telomeres and rDNA in the sir2 mutant strains. (A) Cross-linked chromatin was immunoprecipitated with antibodies to Sir3p. To test for localization of the Sir complex at the telomeres, the DNA pulled down was probed for telomere sequences by using PCR primers corresponding to 300 base pairs from the end of the telomere at the right arm of chromosome VI. Threefold dilutions of the immunoprecipitated DNA were used in the PCR reaction. (B) Pulled down DNA was probed for sequences corresponding to an internal nonsilenced location on chromosome XII. (C) Pulled down DNA from wild type, Δsir2, and mutant 270 was probed for more distal telomere sequences by using PCR primers that recognize sequences that are 3000 base pairs from the end of the telomere at the right arm of chromosome VI. (D) Cross-linked chromatin pulled down with anti-Sir2p was blotted to a membrane by using a slot blotting apparatus and probed for rDNA sequences.
References
-
- Aparicio OM, Billington BL, Gottschling DE. Modifiers of position effect are shared between telomeric and silent mating-type loci in S. cerevisiae. Cell. 1991;66:1279–1287. - PubMed
-
- Braunstein M, Rose AB, Holmes SG, Allis CD, Broach JR. Transcriptional silencing in yeast is associated with reduced nucleosome acetylation. Genes Dev. 1993;7:592–604. - PubMed
-
- Bryk M, Banerjee M, Murphy M, Knudsen KE, Garfinkel DJ, Curcio MJ. Transcriptional silencing of Ty1 elements in the RDN1 locus of yeast. Genes Dev. 1997;11:255–269. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
