

Conditional switching of VEGF provides new insights into adult neovascularization and pro-angiogenic therapy
- PMID: 11953313
- PMCID: PMC125962
- DOI: 10.1093/emboj/21.8.1939
Conditional switching of VEGF provides new insights into adult neovascularization and pro-angiogenic therapy
Abstract
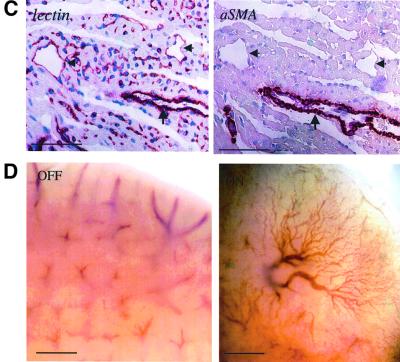
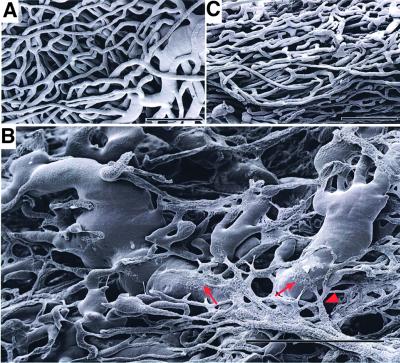
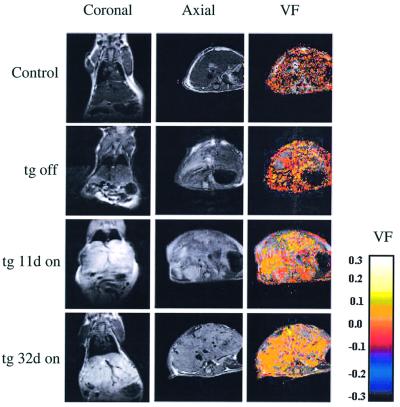
To gain insight into neovascularization of adult organs and to uncover inherent obstacles in vascular endothelial growth factor (VEGF)-based therapeutic angiogenesis, a transgenic system for conditional switching of VEGF expression was devised. The system allows for a reversible induction of VEGF specifically in the heart muscle or liver at any selected schedule, thereby circumventing embryonic lethality due to developmental misexpression of VEGF. Using this system, we demonstrate a progressive, unlimited ramification of the existing vasculature. In the absence of spatial cues, however, abnormal vascular trees were produced, a consequence of chaotic connections with the existing network and formation of irregularly shaped sac-like vessels. VEGF also caused a massive and highly disruptive edema. Importantly, premature cessation of the VEGF stimulus led to regression of most acquired vessels, thus challenging the utility of therapeutic approaches relying on short stimulus duration. A critical transition point was defined beyond which remodeled new vessels persisted for months after withdrawing VEGF, conferring a long-term improvement in organ perfusion. This novel genetic system thus highlights remaining problems in the implementation of pro-angiogenic therapy.
Figures





References
-
- Abramovitch R., Frenkiel,D. and Neeman,M. (1998) Analysis of subcutaneous angiogenesis by gradient echo magnetic resonance imaging. Magn. Reson. Med., 39, 813–824. - PubMed
-
- Adams R.H. and Klein,R. (2000) Eph receptors and ephrin ligands. Essential mediators of vascular development. Trends Cardiovasc. Med., 10, 183–188. - PubMed
-
- Alon T., Hemo,I., Itin,A., Pe’er,J., Stone,J. and Keshet,E. (1995) Vascular endothelial growth factor acts as a survival factor for newly formed retinal vessels and has implications for retinopathy of prematurity. Nature Med., 1, 1024–1028. - PubMed
-
- Benjamin L.E., Hemo,I. and Keshet,E. (1998) A plasticity window for blood vessel remodelling is defined by pericyte coverage of the preformed endothelial network and is regulated by PDGF-B and VEGF. Development, 125, 1591–1598. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
