

Global analysis of outer membrane proteins from Leptospira interrogans serovar Lai
- PMID: 11953365
- PMCID: PMC127947
- DOI: 10.1128/IAI.70.5.2311-2318.2002
Global analysis of outer membrane proteins from Leptospira interrogans serovar Lai
Abstract
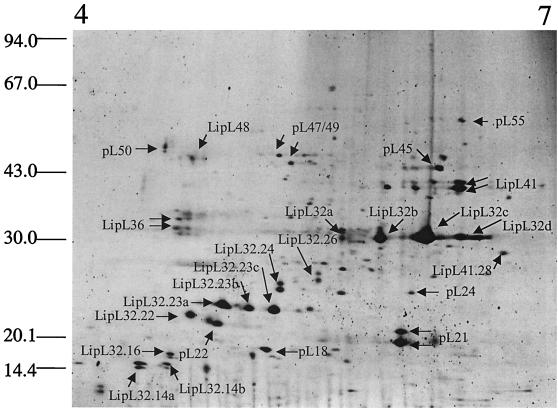
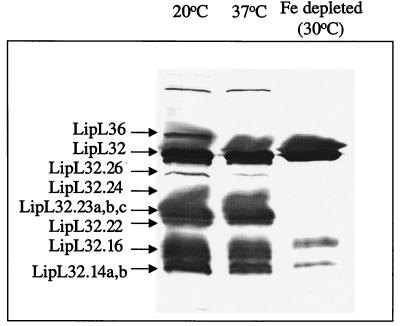
Recombinant leptospiral outer membrane proteins (OMPs) can elicit immunity to leptospirosis in a hamster infection model. Previously characterized OMPs appear highly conserved, and thus their potential to stimulate heterologous immunity is of critical importance. In this study we undertook a global analysis of leptospiral OMPs, which were obtained by Triton X-114 extraction and phase partitioning. Outer membrane fractions were isolated from Leptospira interrogans serovar Lai grown at 20, 30, and 37 degrees C with or without 10% fetal calf serum and, finally, in iron-depleted medium. The OMPs were separated by two-dimensional gel electrophoresis. Gel patterns from each of the five conditions were compared via image analysis, and 37 gel-purified proteins were tryptically digested and characterized by mass spectrometry (MS). Matrix-assisted laser desorption ionization-time-of-flight MS was used to rapidly identify leptospiral OMPs present in sequence databases. Proteins identified by this approach included the outer membrane lipoproteins LipL32, LipL36, LipL41, and LipL48. No known proteins from any cellular location other than the outer membrane were identified. Tandem electrospray MS was used to obtain peptide sequence information from eight novel proteins designated pL18, pL21, pL22, pL24, pL45, pL47/49, pL50, and pL55. The expression of LipL36 and pL50 was not apparent at temperatures above 30 degrees C or under iron-depleted conditions. The expression of pL24 was also downregulated after iron depletion. The leptospiral major OMP LipL32 was observed to undergo substantial cleavage under all conditions except iron depletion. Additionally, significant downregulation of these mass forms was observed under iron limitation at 30 degrees C, but not at 30 degrees C alone, suggesting that LipL32 processing is dependent on iron-regulated extracellular proteases. However, separate cleavage products responded differently to changes in growth temperature and medium constituents, indicating that more than one process may be involved in LipL32 processing. Furthermore, under iron-depleted conditions there was no concomitant increase in the levels of the intact form of LipL32. The temperature- and iron-regulated expression of LipL36 and the iron-dependent cleavage of LipL32 were confirmed by immunoblotting with specific antisera. Global analysis of the cellular location and expression of leptospiral proteins will be useful in the annotation of genomic sequence data and in providing insight into the biology of Leptospira.
Figures


References
-
- Altschul, S. F., G. Warren, W. Miller, E. Myers, and D. J. Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403-410. - PubMed
-
- Branger, C., C. Sonrier, B. Chatrenet, B. Klonjkowski, N. Ruvoen-Clouet, A. Aubert, G. Andre-Fontaine, and M. Eloit. 2001. Identification of the hemolysis-associated protein 1 as a cross-protective immunogen of Leptospira interrogans by adenovirus-mediated vaccination. Infect. Immun. 69:6831-6838. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
