

SETDB1: a novel KAP-1-associated histone H3, lysine 9-specific methyltransferase that contributes to HP1-mediated silencing of euchromatic genes by KRAB zinc-finger proteins
- PMID: 11959841
- PMCID: PMC152359
- DOI: 10.1101/gad.973302
SETDB1: a novel KAP-1-associated histone H3, lysine 9-specific methyltransferase that contributes to HP1-mediated silencing of euchromatic genes by KRAB zinc-finger proteins
Abstract
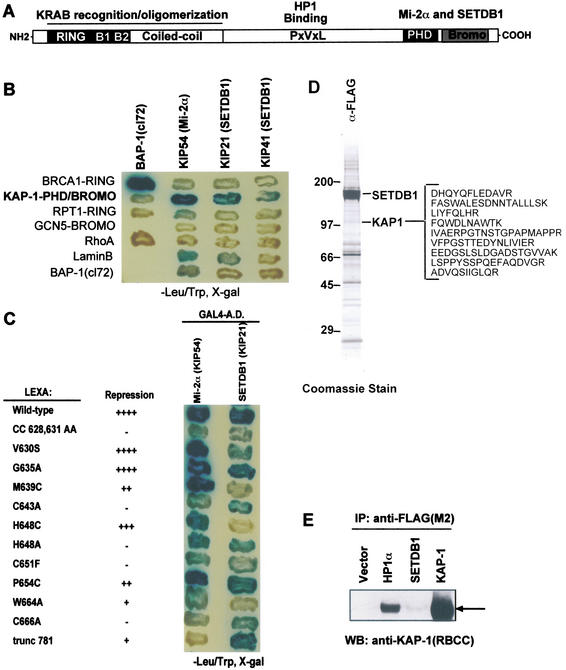
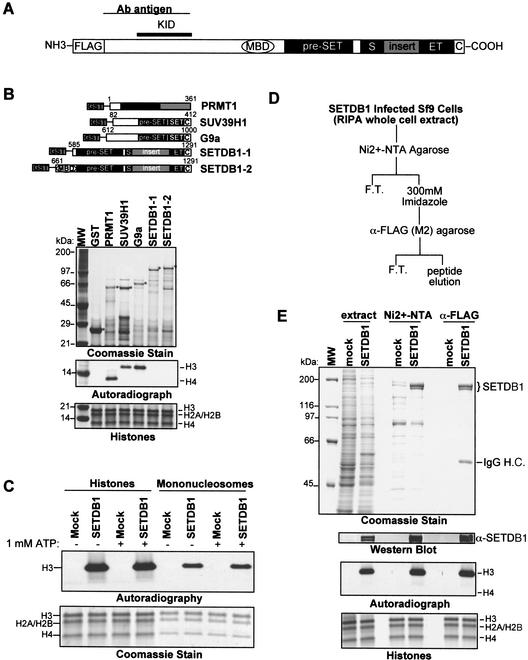
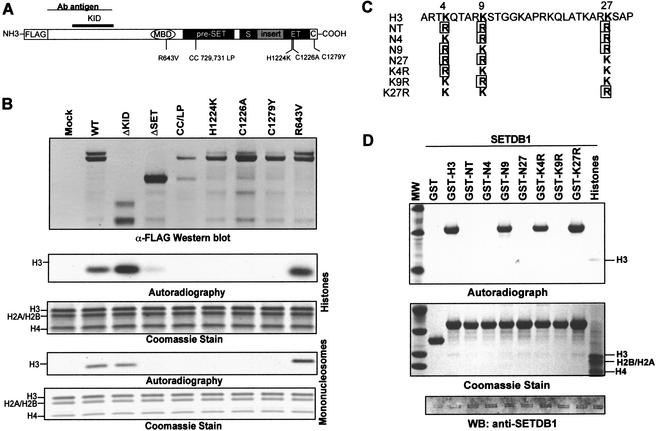
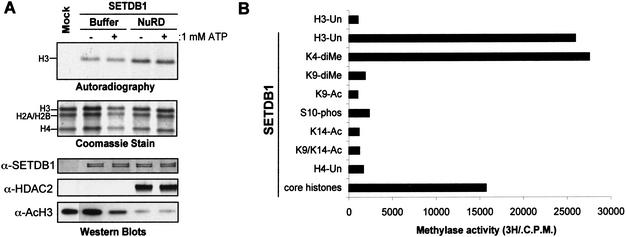
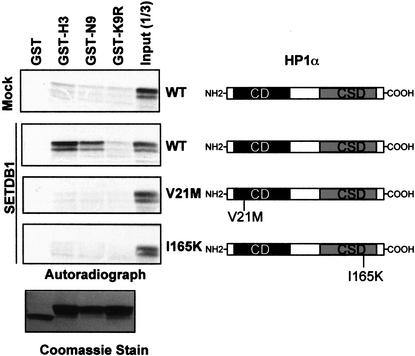
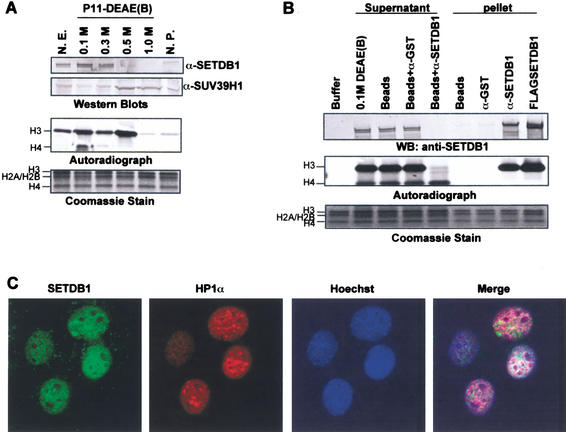
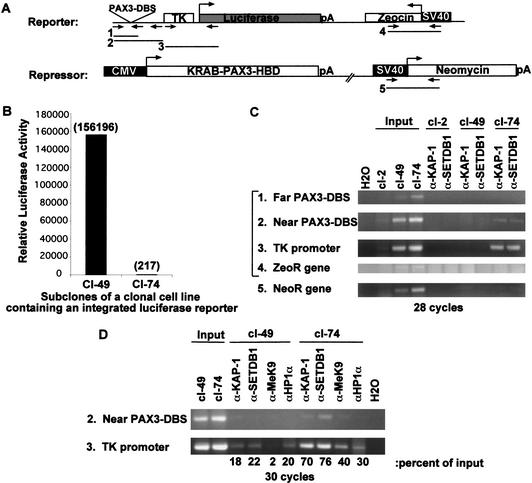
Posttranslational modification of histones has emerged as a key regulatory signal in eukaryotic gene expression. Recent genetic and biochemical studies link H3-lysine 9 (H3-K9) methylation to HP1-mediated heterochromatin formation and gene silencing. However, the mechanisms that target and coordinate these activities to specific genes is poorly understood. Here we report that the KAP-1 corepressor for the KRAB-ZFP superfamily of transcriptional silencers binds to SETDB1, a novel SET domain protein with histone H3-K9-specific methyltransferase activity. Although acetylation and phosphorylation of the H3 N-terminal tail profoundly affect the efficiency of H3-K9 methylation by SETDB1, we found that methylation of H3-K4 does not affect SETDB1-mediated methylation of H3-K9. In vitro methylation of the N-terminal tail of histone H3 by SETDB1 is sufficient to enhance the binding of HP1 proteins, which requires both an intact chromodomain and chromoshadow domain. Indirect immunofluoresence staining of interphase nuclei localized SETDB1 predominantly in euchromatic regions that overlap with HP1 staining in nonpericentromeric regions of chromatin. Moreover, KAP-1, SETDB1, H3-MeK9, and HP1 are enriched at promoter sequences of a euchromatic gene silenced by the KRAB-KAP-1 repression system. Thus, KAP-1 is a molecular scaffold that is targeted by KRAB-ZFPs to specific loci and coordinates both histone methylation and the deposition of HP1 proteins to silence gene expression.
Figures
References
-
- Ayyanathan K, Fredericks WJ, Berking C, Herlyn M, Balakrishnan C, Gunther E, Rauscher FJ., III Hormone-dependent tumor regression in vivo by an inducible transcriptional repressor directed at the PAX3-FKHR oncogene. Cancer Res. 2000;60:5803–5814. - PubMed
-
- Bannister AJ, Zegerman P, Partridge JF, Miska EA, Thomas JO, Allshire RC, Kouzarides T. Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature. 2001;410:120–124. - PubMed
-
- Berger SL. An embarrassment of niches: The many covalent modifications of histones in transcriptional regulation. Oncogene. 2001;20:3007–3013. - PubMed
-
- Boggs BA, Cheung P, Heard E, Spector DL, Chinault AC, Allis CD. Differentially methylated forms of histone H3 show unique association patterns with inactive human X chromosomes. Nat Genet. 2002;30:73–76. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous