

Knotting probability of DNA molecules confined in restricted volumes: DNA knotting in phage capsids
- PMID: 11959991
- PMCID: PMC122776
- DOI: 10.1073/pnas.032095099
Knotting probability of DNA molecules confined in restricted volumes: DNA knotting in phage capsids
Abstract
When linear double-stranded DNA is packed inside bacteriophage capsids, it becomes highly compacted. However, the phage is believed to be fully effective only if the DNA is not entangled. Nevertheless, when DNA is extracted from a tailless mutant of the P4 phage, DNA is found to be cyclic and knotted (probability of 0.95). The knot spectrum is very complex, and most of the knots have a large number of crossings. We quantified the frequency and crossing numbers of these knots and concluded that, for the P4 tailless mutant, at least half the knotted molecules are formed while the DNA is still inside the viral capsid rather than during extraction. To analyze the origin of the knots formed inside the capsid, we compared our experimental results to Monte Carlo simulations of random knotting of equilateral polygons in confined volumes. These simulations showed that confinement of closed chains to tightly restricted volumes results in high knotting probabilities and the formation of knots with large crossing numbers. We conclude that the formation of the knots inside the viral capsid is driven mainly by the effects of confinement.
Figures

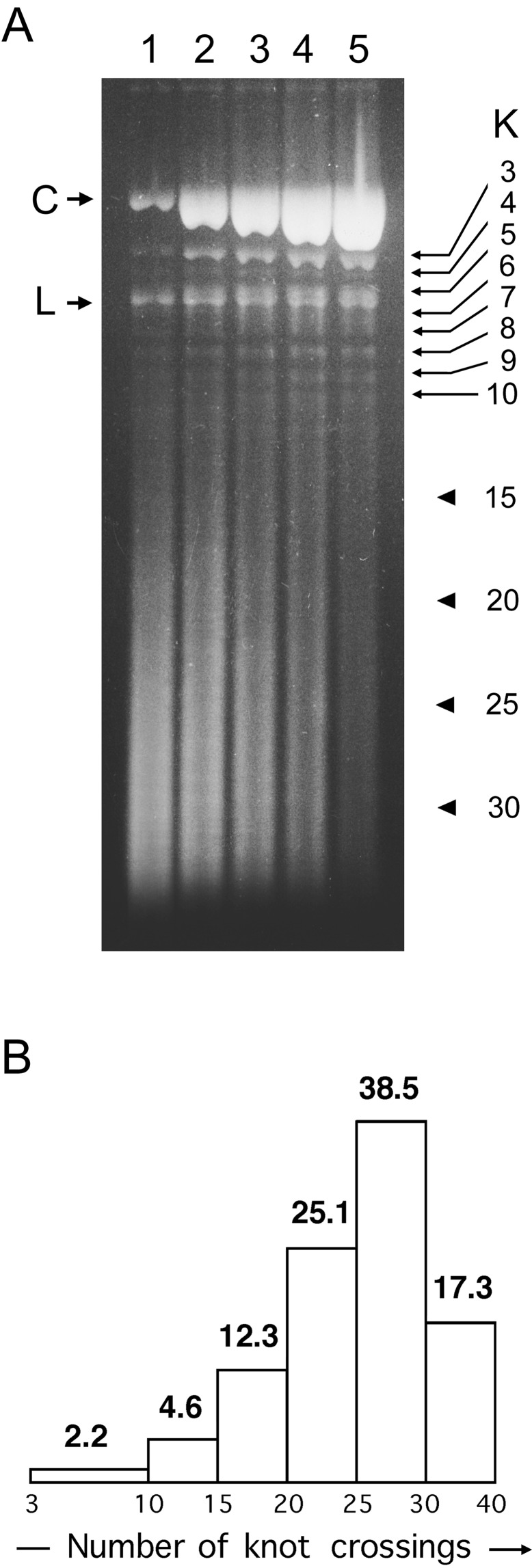
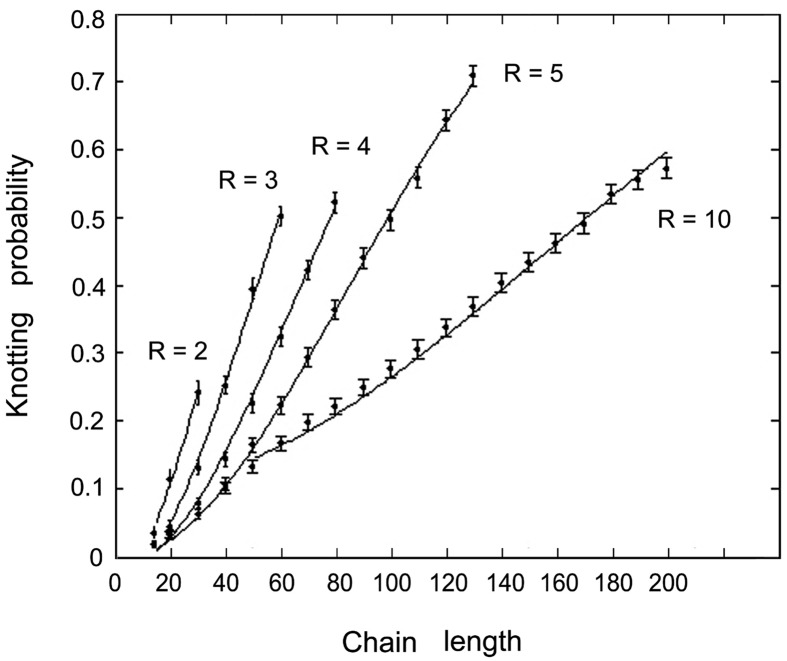

References
-
- Shaw S Y, Wang J C. Science. 1993;260:533–536. - PubMed
-
- Wasserman S A, Cozzarelli N R. Science. 1986;232:951–960. - PubMed
-
- Stark W M, Boocock M R, Sherratt D J. Trends Genet. 1989;5:304–309. - PubMed
-
- Frank-Kamentskii M D, Lukashin A V, Anshelevich V V, Vologodskii A V. J Biomol Struct Dyn. 1985;2:1005–1012. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
