

Developmental specificity of recruitment of TBP to the TATA box of the human gamma-globin gene
- PMID: 11960008
- PMCID: PMC122800
- DOI: 10.1073/pnas.072084499
Developmental specificity of recruitment of TBP to the TATA box of the human gamma-globin gene
Abstract
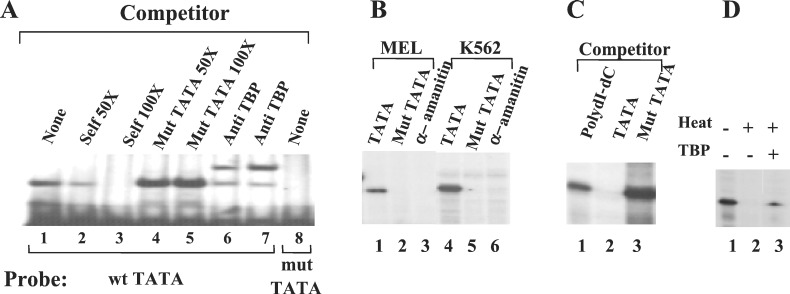
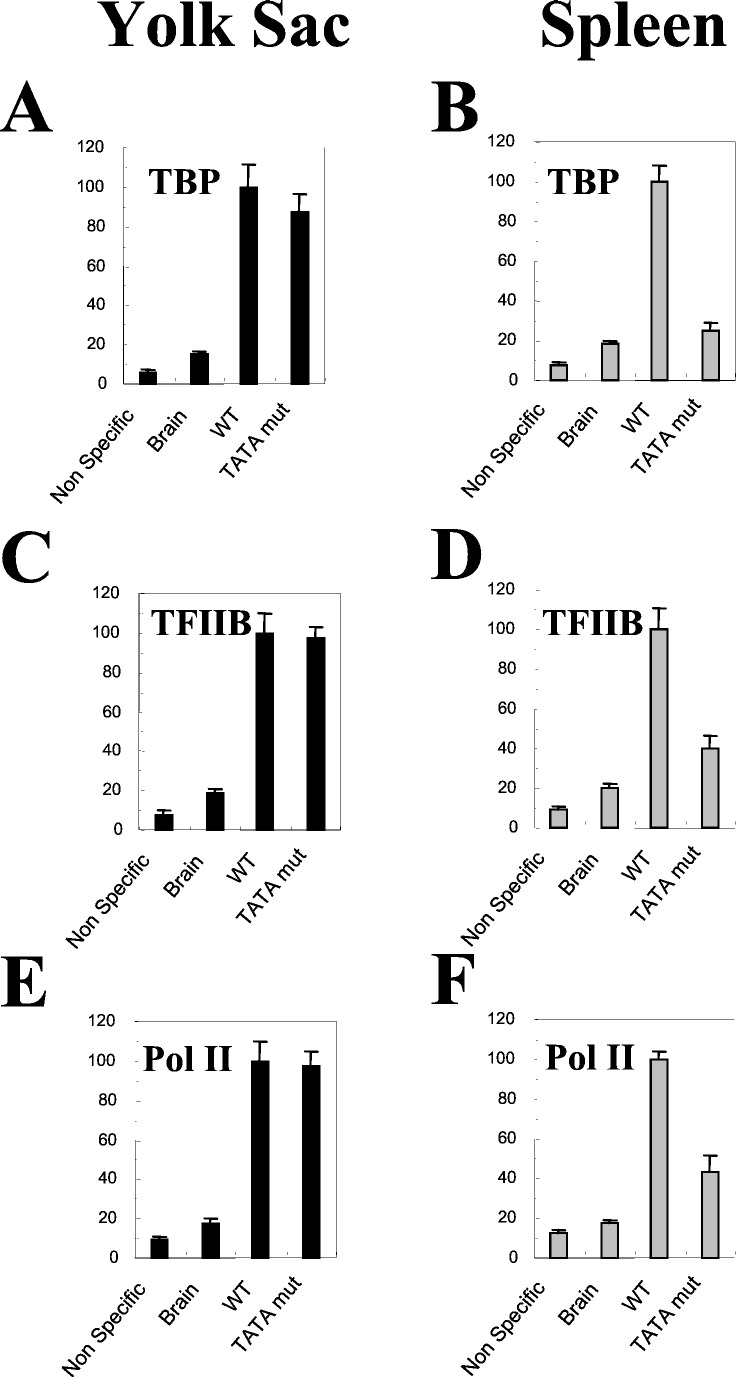
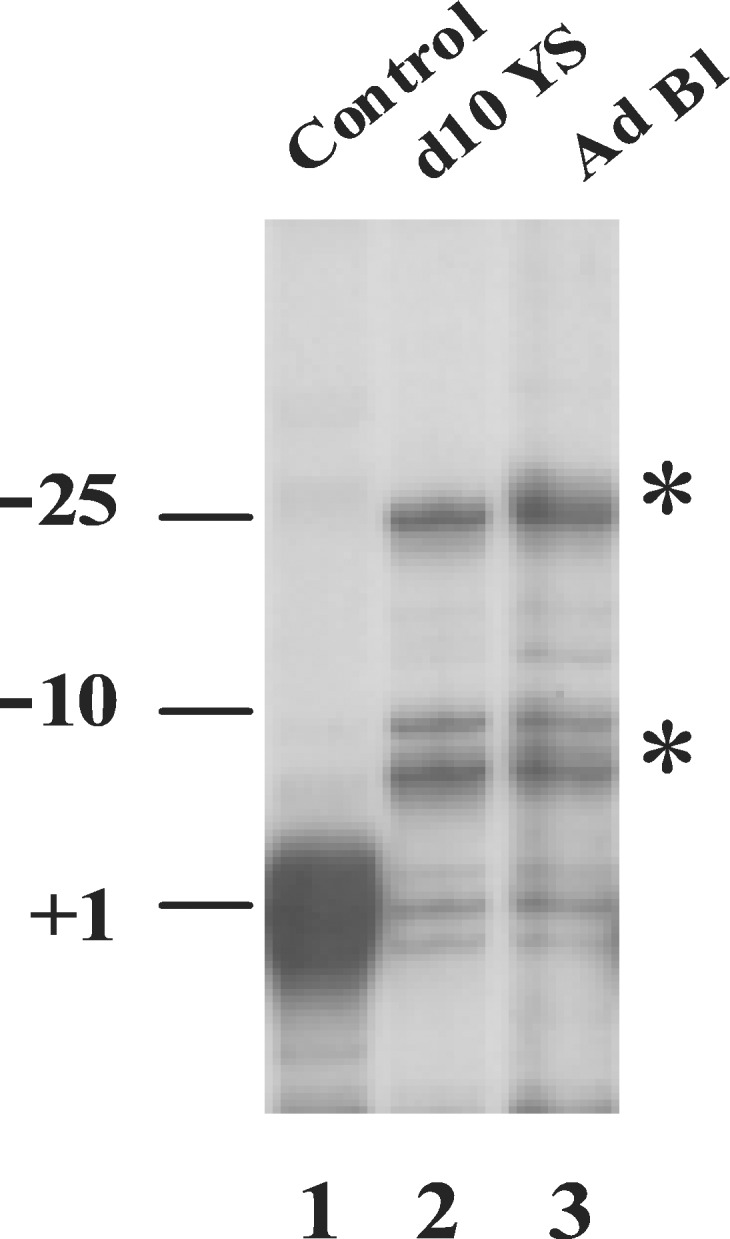
It is unclear whether the core promoter is involved in developmental regulation. To address this question, we mutated the TATA box of the human gamma-globin gene, produced transgenic mice, and examined the effect of the mutation during the course of mouse development. In our test system, the gamma-globin gene is expressed at similar levels in the embryonic and adult erythroid cells. The TATA box mutation dramatically reduced expression of the gamma-globin gene in the adult but not in embryonic erythroid cells. In addition, the disruption of the gamma TATA box significantly reduced the recruitment of TATA box-binding protein (TBP) in the adult cells, but not in embryonic cells, suggesting that the recruitment of TBP to the gamma gene promoter is developmentally specific. Similarly, the recruitment of transcription factor II B and RNA polymerase II to the gamma promoter was affected in the adult but not in embryonic cells. The distinct effects of the TATA mutation in the embryonic and adult developmental stages suggest that the basal transcription apparatus can be recruited to a core promoter in a developmental stage-dependent manner. The TATA mutation resulted in a shift of transcription initiation site 6 bp or longer upstream to the cap site both in the embryonic and adult erythrocytes. We conclude that the TATA box determines the initiation site but not the efficiency of transcription of the gamma-globin gene.
Figures
References
-
- Holmes M C, Tjian R. Science. 2000;288:867–870. - PubMed
-
- Kuras L, Kosa P, Mencia M, Struhl K. Science. 2000;288:1244–1248. - PubMed
-
- Li X Y, Bhaumik S R, Green M R. Science. 2000;288:1242–1244. - PubMed
-
- Dantonel J C, Quintin S, Lakatos L, Labouesse M, Tora L. Mol Cell. 2000;6:715–722. - PubMed
-
- Kaltenbach L, Horner M A, Rothman J H, Mango S E. Mol Cell. 2000;6:705–713. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
