

Holding two heads together: stability of the myosin II rod measured by resonance energy transfer between the heads
- PMID: 11972024
- PMCID: PMC122893
- DOI: 10.1073/pnas.082024299
Holding two heads together: stability of the myosin II rod measured by resonance energy transfer between the heads
Abstract
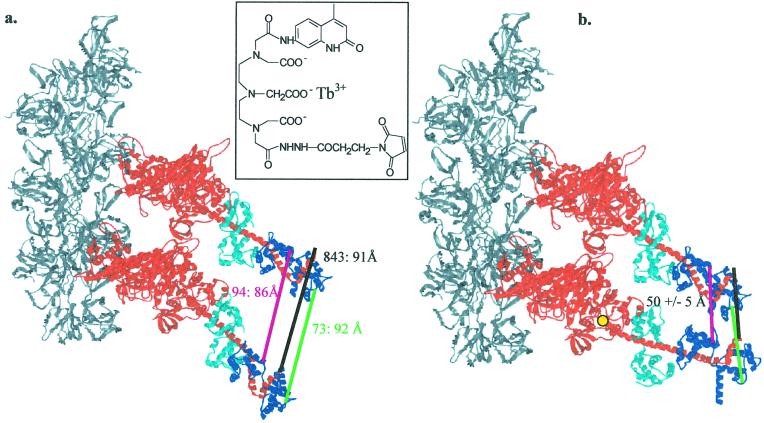
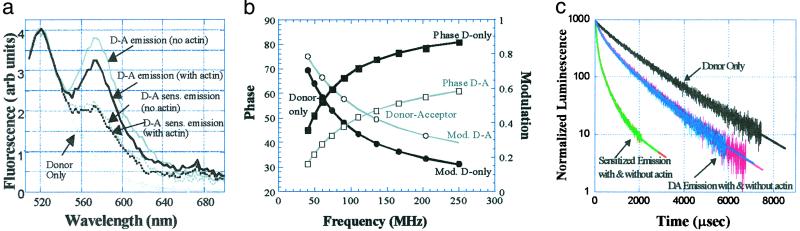
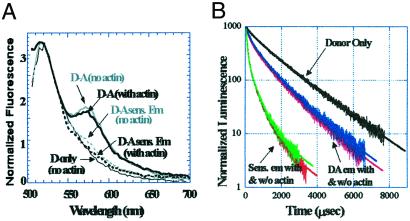
Myosin, similar to many molecular motors, is a two-headed dimer held together by a coiled-coiled rod. The stability of the coiled coil has implications for head-head interactions, force generation, and possibly regulation. Here we used two different resonance energy transfer techniques to measure the distances between probes placed in the regulatory light chain of each head of a skeletal heavy meromyosin, near the head-rod junction (positions 2, 73, and 94). Our results indicate that the rod largely does not uncoil when myosin is free in solution, and at least beyond the first heptad, the subfragment 2 rod remains relatively intact even under the relatively large strain of two-headed myosin (rigor) binding to actin. We infer that uncoiling of the rod likely does not play a role in myosin II motility. To keep the head-rod junction intact, a distortion must occur within the myosin heads. This distortion may lead to different orientations of the light-chain domains within the myosin dimer when both heads are attached to actin, which would explain previously puzzling observations and require reinterpretation of others. In addition, by comparing resonance energy transfer techniques sensitive to different dynamical time scales, we find that the N terminus of the regulatory light chain is highly flexible, with possible implications for regulation. An intact rod may be a general property of molecular motors, because a similar conclusion has been reached recently for kinesin, although whether the rod remains intact will depend on the relative stiffness of the coiled coil and the head in different motors.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
