

Naturally occurring rhodopsin mutation in the dog causes retinal dysfunction and degeneration mimicking human dominant retinitis pigmentosa
- PMID: 11972042
- PMCID: PMC122948
- DOI: 10.1073/pnas.082714499
Naturally occurring rhodopsin mutation in the dog causes retinal dysfunction and degeneration mimicking human dominant retinitis pigmentosa
Abstract
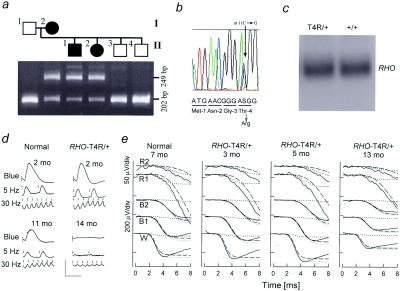
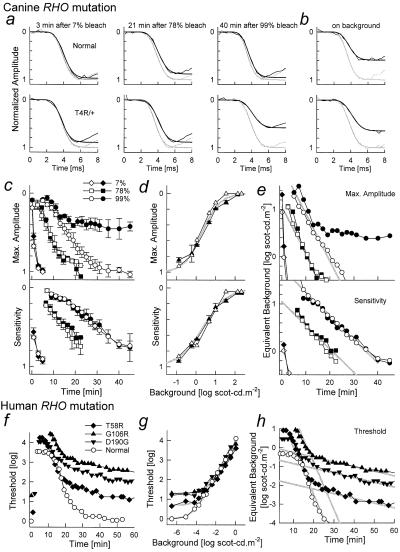
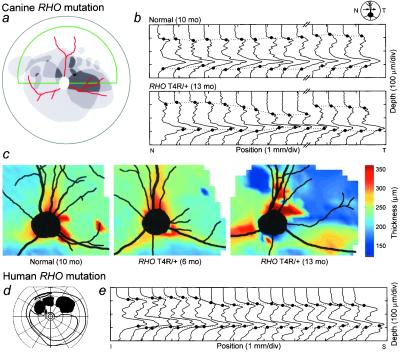
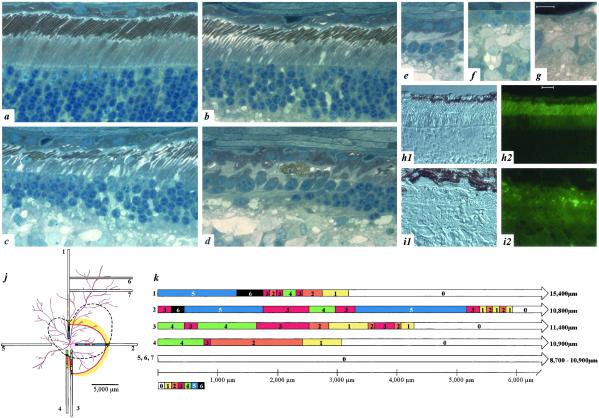
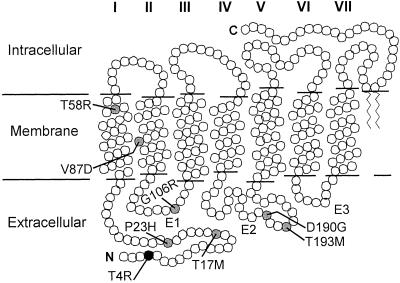
Rhodopsin is the G protein-coupled receptor that is activated by light and initiates the transduction cascade leading to night (rod) vision. Naturally occurring pathogenic rhodopsin (RHO) mutations have been previously identified only in humans and are a common cause of dominantly inherited blindness from retinal degeneration. We identified English Mastiff dogs with a naturally occurring dominant retinal degeneration and determined the cause to be a point mutation in the RHO gene (Thr4Arg). Dogs with this mutant allele manifest a retinal phenotype that closely mimics that in humans with RHO mutations. The phenotypic features shared by dog and man include a dramatically slowed time course of recovery of rod photoreceptor function after light exposure and a distinctive topographic pattern to the retinal degeneration. The canine disease offers opportunities to explore the basis of prolonged photoreceptor recovery after light in RHO mutations and determine whether there are links between the dysfunction and apoptotic retinal cell death. The RHO mutant dog also becomes the large animal needed for preclinical trials of therapies for a major subset of human retinopathies.
Figures
References
-
- Spiegel A M. Annu Rev Physiol. 1995;58:143–170. - PubMed
-
- Rattner A, Sun H, Nathans J. Annu Rev Genet. 1999;33:89–131. - PubMed
-
- Palczewski K, Kumasaka T, Hori T, Behnke C A, Motoshima H, Fox B A, Le Trong I, Teller D C, Okada T, Stenkamp R E, et al. Science. 2000;289:739–745. - PubMed
-
- Gal A, Apfelstedt-Sylla E, Janecke A R, Zrenner E. Prog Ret Eye Res. 1997;16:51–79.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
