

Molecular and pharmacological characterization of genes encoding urotensin-II peptides and their cognate G-protein-coupled receptors from the mouse and monkey
- PMID: 11976263
- PMCID: PMC1762106
- DOI: 10.1038/sj.bjp.0704671
Molecular and pharmacological characterization of genes encoding urotensin-II peptides and their cognate G-protein-coupled receptors from the mouse and monkey
Abstract
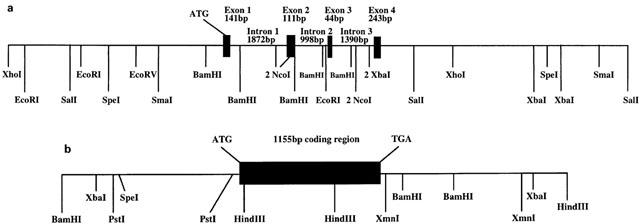
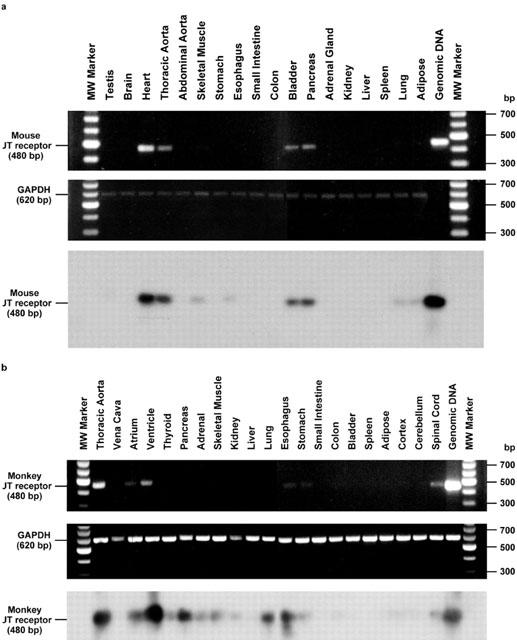
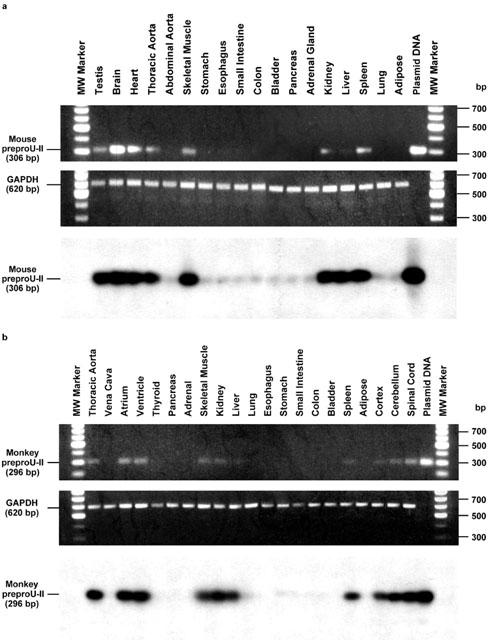
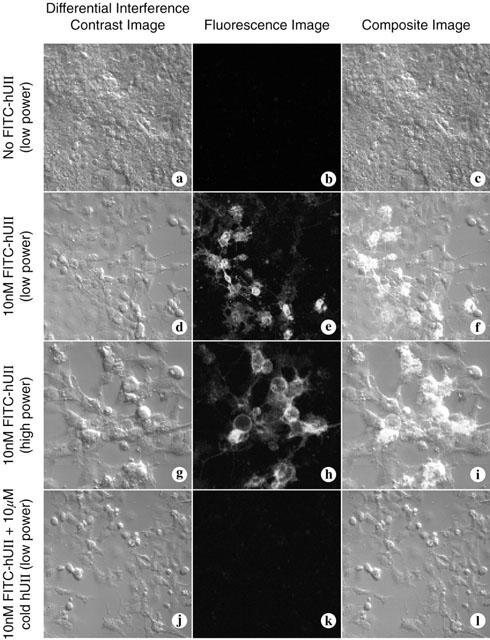
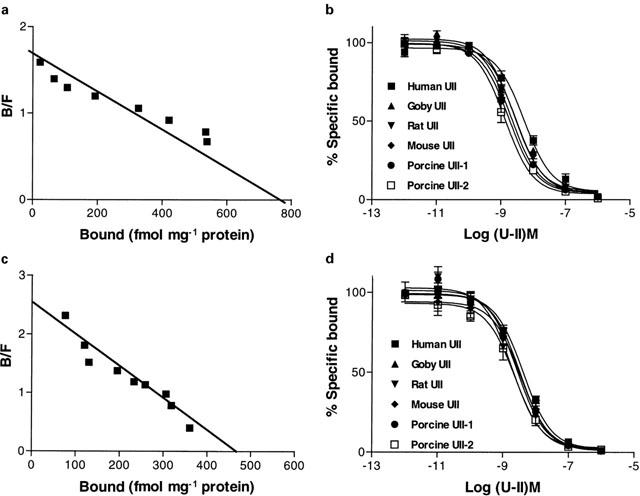
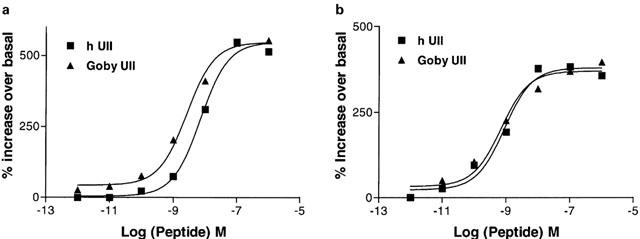
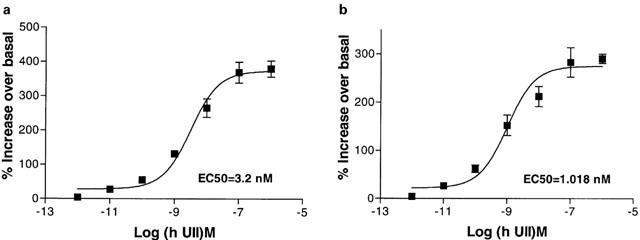
Urotensin-II (U-II) and its receptor (UT) represent novel therapeutic targets for management of a variety of cardiovascular diseases. To test such hypothesis, it will be necessary to develop experimental animal models for the manipulation of U-II/UT receptor system. The goal of this study was to clone mouse and primate preproU-II and UT for pharmacological profiling. Monkey and mouse preproU-II genes were identified to encode 123 and 125 amino acids. Monkey and mouse UT receptors were 389, and 386 amino acids, respectively. Genomic organization of mouse genes showed that the preproU-II has four exons, while the UT receptor has one exon. Although initially viewed by many exclusively as cardiovascular targets, the present study demonstrates expression of mouse and monkey U-II/UT receptor mRNA in extra-vascular tissue including lung, pancreas, skeletal muscle, kidney and liver. Ligand binding studies showed that [125I]h U-II bound to a single sites to the cloned receptors in a saturable/high affinity manner (Kd 654+/-154 and 214+/-65 pM and Bmax of 1011+/-125 and 497+/-68 fmol mg-1 for mouse and monkey UT receptors, respectively). Competition binding analysis demonstrated equipotent, high affinity binding of numerous mammalian, amphibian and piscine U-II isopeptides to these receptors (Ki=0.8 - 3 nM). Fluorescein isothiocyanate (FITC) labelled U-II, bound specifically to HEK-293 cells expressing mouse or monkey UT receptor, confirming cell surface expression of recombinant UT receptor. Exposure of these cells to human U-II resulted in an increase in intracellular [Ca2+] concentrations (EC50 3.2+/-0.8 and 1.1+/-0.3 nM for mouse and monkey UT receptors, respectively) and inositol phosphate (Ip) formation (EC50 7.2+/-1.8 and 0.9+/-0.2 nM for mouse and monkey UT receptors, respectively) consistent with the primary signalling pathway for UT receptor involving phospholipase C activation.
Figures


References
-
- AIYAR N., BAKER E., WU H.L., NAMBI P., EDWARDS R.M., TRILL J.J., ELLIS C., BERGSMA D.J. Human AT1 receptor is a single copy gene: characterization in a stable cell line. Mol. Cell Biochem. 1994;131:75–86. - PubMed
-
- AIYAR N.V., NAMBI P., STASSEN F.L., CROOKE S.T. Vascular vasopressin receptors mediate phosphatidylinositol turnover and calcium efflux in an established smooth muscle cell line. Life Sciences. 1986;39:37–45. - PubMed
-
- AMES R.S., SARAU H.M., CHAMBERS J.K., WILLETTE R.N., AIYAR N.V., ROMANIC A.M., LOUDEN C.S., FOLEY J.J., SAUERMELCH C.F., COATNEY R.W., AO Z., DISA J., HOLMES S.D., STADEL J.M., MARTIN J.D., LIU W.S., GLOVER G.I., WILSON S., MCNULTY D.E., ELLIS C.E., ELSHOURBAGY N.A., SHABON U., TRILL J.J., HAY D.W., OHLSTEIN E.H., BERGSMA D.J., DOUGLAS S.A. Human urotensin-II is a potent vasoconstrictor and agonist for the orphan receptor GPR14. Nature. 1999;401:282–286. - PubMed
-
- BERN H.A., LEDERIS K. A reference preparation for the study of active substances in the caudal neurosecretory system of teleosts. J. Endocrinol. 1969;45:xi–xii. - PubMed
-
- BERN H.A., PEARSON D., LARSON B.A., NISHIOKA R.S. Neurohormones from fish tails: the caudal neurosecretory system. I. ‘Urophysiology' and the caudal neurosecretory system of fishes. Rec. Prog. Horm. Res. 1985;41:533–552. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
