

Export of autotransported proteins proceeds through an oligomeric ring shaped by C-terminal domains
- PMID: 11980709
- PMCID: PMC125980
- DOI: 10.1093/emboj/21.9.2122
Export of autotransported proteins proceeds through an oligomeric ring shaped by C-terminal domains
Abstract
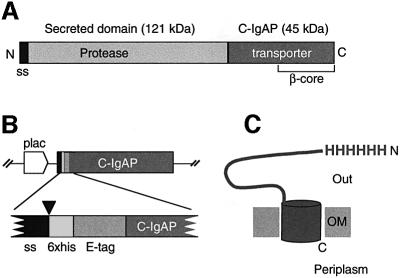
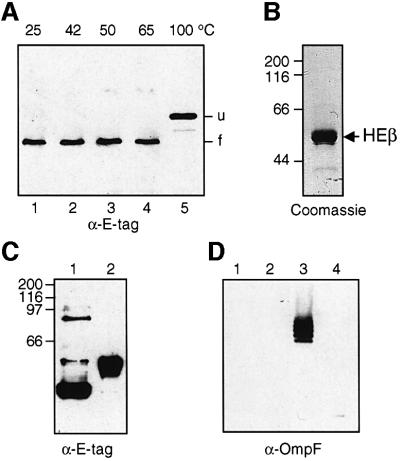
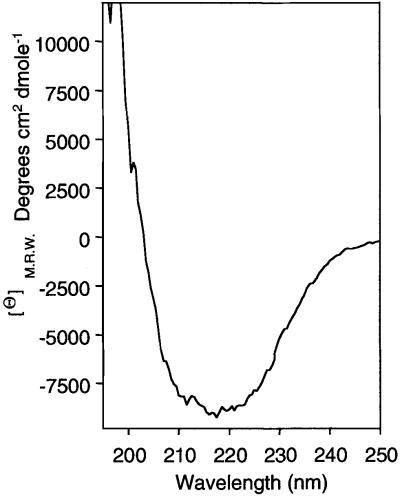
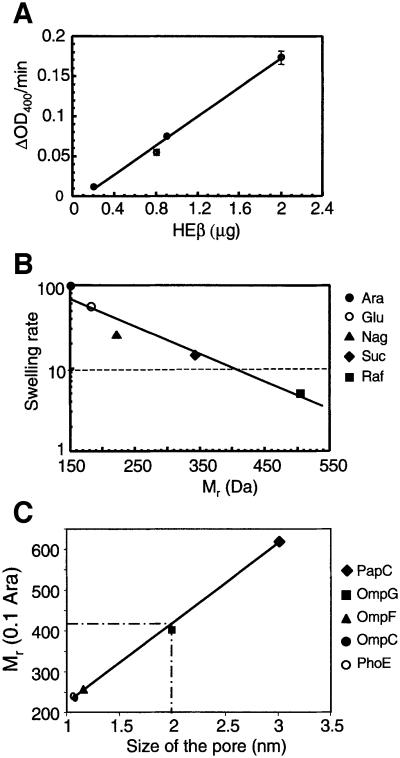
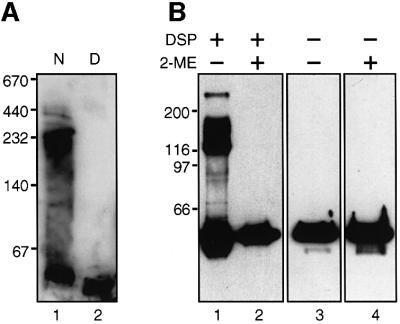
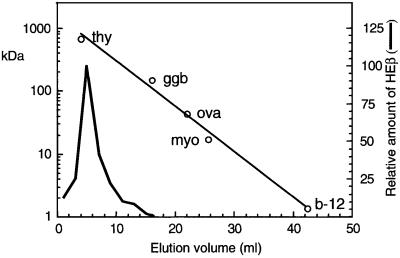
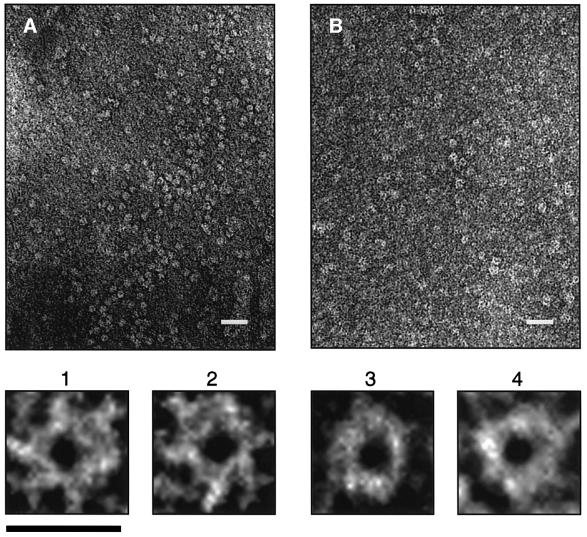
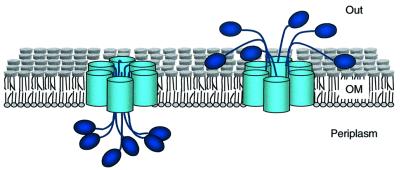
An investigation was made into the oligomerization, the ability to form pores and the secretion-related properties of the 45 kDa C-terminal domain of the IgA protease (C-IgAP) from Neisseria gonorrhoeae. This protease is the best studied example of the autotransporters (ATs), a large family of exoproteins from Gram-negative bacteria that includes numerous virulence factors from human pathogens. These proteins contain an N-terminal passenger domain that em bodies the secreted polypeptide, while the C-domain inserts into the outer membrane (OM) and trans locates the linked N-module into the extracellular medium. Here we report that purified C-IgAP forms an oligomeric complex of approximately 500 kDa with a ring-like structure containing a central cavity of approximately 2 nm diameter that is the conduit for the export of the N-domains. These data overcome the previous model for ATs, which postulated the passage of the N-module through the hydrophilic channel of the beta-barrel of each monomeric C-domain. Our results advocate a secretion mechanism not unlike other bacterial export systems, such as the secretins or fimbrial ushers, which rely on multimeric complexes assembled in the OM.
Figures
References
-
- Ausubel F.M., Brent,R., Kingston,R.E., Moore,D.D., Seidman,J.G., Smith,J.A. and Struhl,K. (1994) Current Protocols in Molecular Biology. John Wiley & Sons, New York, NY.
-
- Ay J., Keitel,T., Kuttner,G., Wessner,H., Scholz,C., Hahn,M. and Hohne,W. (2000) Crystal structure of a phage library-derived single-chain Fv fragment complexed with turkey egg-white lysozyme at 2.0 Å resolution. J. Mol. Biol., 301, 239–246. - PubMed
-
- Benz I. and Schmidt,M.A. (1992) AIDA-I, the adhesin involved in diffuse adherence of diarrhoeagenic Escherichia coli strain 2787 (O126:H27), is synthesized via a precursor molecule. Mol. Microbiol., 6, 1539–1546. - PubMed
-
- Bernstein H.D. (2000) The biogenesis and assembly of bacterial membrane proteins. Curr. Opin. Microbiol., 3, 203–209. - PubMed
-
- Bitter W., Koster,M., Latijnhouwers,M., de Cock,H. and Tommassen,J. (1998) Formation of oligomeric rings by XcpQ and PilQ, which are involved in protein transport across the outer membrane of Pseudomonas aeruginosa. Mol. Microbiol., 27, 209–219. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
