

ATP-dependent DNA ligases
- PMID: 11983065
- PMCID: PMC139351
- DOI: 10.1186/gb-2002-3-4-reviews3005
ATP-dependent DNA ligases
Abstract
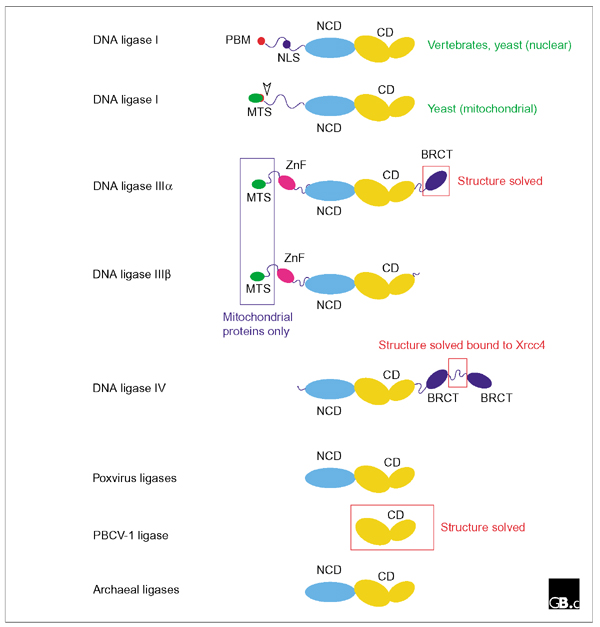
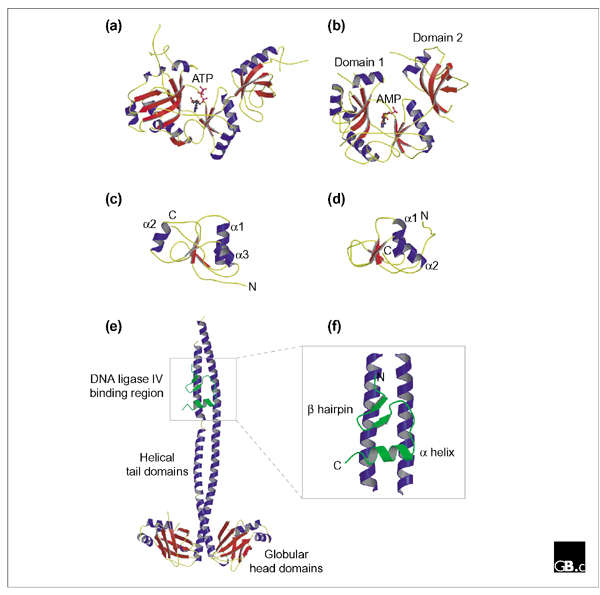
By catalyzing the joining of breaks in the phosphodiester backbone of duplex DNA, DNA ligases play a vital role in the diverse processes of DNA replication, recombination and repair. Three related classes of ATP-dependent DNA ligase are readily apparent in eukaryotic cells. Enzymes of each class comprise catalytic and non-catalytic domains together with additional domains of varying function. DNA ligase I is required for the ligation of Okazaki fragments during lagging-strand DNA synthesis, as well as for several DNA-repair pathways; these functions are mediated, at least in part, by interactions between DNA ligase I and the sliding-clamp protein PCNA. DNA ligase III, which is unique to vertebrates, functions both in the nucleus and in mitochondria. Two distinct isoforms of this enzyme, differing in their carboxy-terminal sequences, are produced by alternative splicing: DNA ligase IIIalpha has a carboxy-terminal BRCT domain that interacts with the mammalian DNA-repair factor XrccI, but both alpha and beta isoforms have an amino-terminal zinc-finger motif that appears to play a role in the recognition of DNA secondary structures that resemble intermediates in DNA metabolism. DNA ligase IV is required for DNA non-homologous end joining pathways, including recombination of the V(D)J immunoglobulin gene segments in cells of the mammalian immune system. DNA ligase IV forms a tight complex with Xrcc4 through an interaction motif located between a pair of carboxy-terminal BRCT domains in the ligase. Recent structural studies have shed light on the catalytic function of DNA ligases, as well as illuminating protein-protein interactions involving DNA ligases IIIalpha and IV.
Figures
References
-
- Timson DJ, Singleton MR, Wigley DB. DNA ligases in the repair and replication of DNA. Mutat Res. 2000;460:301–318. A detailed overview of DNA ligase structure and function, with particular emphasis on lessons from structural studies. - PubMed
-
- Parks RJ, Winchcombe-Forhan C, DeLange AM, Xing X, Evans DH. DNA ligase gene disruptions can depress viral growth and replication in poxvirus-infected cells. Virus Res. 1998;56:135–147. Functional analysis of the vaccinia and Shope fibroma virus (SFV) ATP-dependent DNA ligases. - PubMed
-
- Shuman S, Schwer B. RNA capping enzyme and DNA ligase: a superfamily of covalent nucleotidyl transferases. Mol Microbiol. 1995;17:405–410. Identification of the nucleotidyl transferase superfamily that includes both DNA ligases (ATP- and NAD+-dependent) as well as GTP-dependent mRNA capping enzymes. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
