

Galactose metabolism is essential for the African sleeping sickness parasite Trypanosoma brucei
- PMID: 11983889
- PMCID: PMC122871
- DOI: 10.1073/pnas.092669999
Galactose metabolism is essential for the African sleeping sickness parasite Trypanosoma brucei
Abstract
The tsetse fly-transmitted protozoan parasite Trypanosoma brucei is the causative agent of human African sleeping sickness and the cattle disease Nagana. The bloodstream form of the parasite uses a dense cell-surface coat of variant surface glycoprotein to escape the innate and adaptive immune responses of the mammalian host and a highly glycosylated transferrin receptor to take up host transferrin, an essential growth factor. These glycoproteins, as well as other flagellar pocket, endosomal, and lysosomal glycoproteins, are known to contain galactose. The parasite is unable to take up galactose, suggesting that it may depend on the action of UDP-glucose 4'-epimerase for the conversion of UDP-Glc to UDP-Gal and subsequent incorporation of galactose into glycoconjugates via UDP-Gal-dependent galactosyltransferases. In this paper, we describe the cloning of T. brucei galE, encoding T. brucei UDP-Glc-4'-epimerase, and functional characterization by complementation of a galE-deficient Escherichia coli mutant and enzymatic assay of recombinant protein. A tetracycline-inducible conditional galE null mutant of T. brucei was created using a transgenic parasite expressing the TETR tetracycline repressor protein gene. Withdrawal of tetracycline led to a cessation of cell division and substantial cell death, demonstrating that galactose metabolism in T. brucei proceeds via UDP-Glc-4'-epimerase and is essential for parasite growth. After several days without tetracycline, cultures spontaneously recovered. These cells were shown to have undergone a genetic rearrangement that deleted the TETR gene. The results show that enzymes and transporters involved in galactose metabolism may be considered as potential therapeutic targets against African trypanosomiasis.
Figures

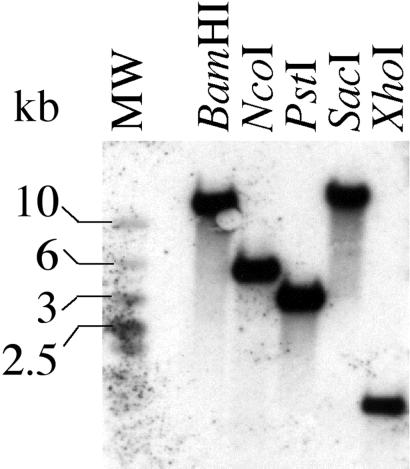
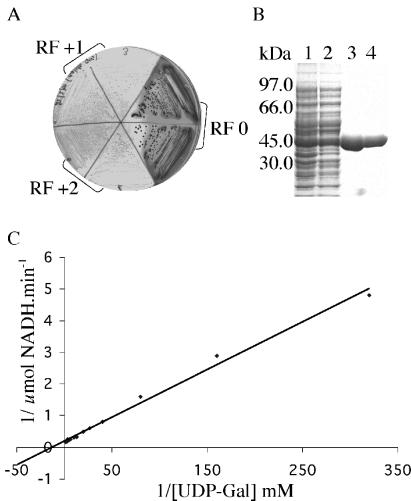
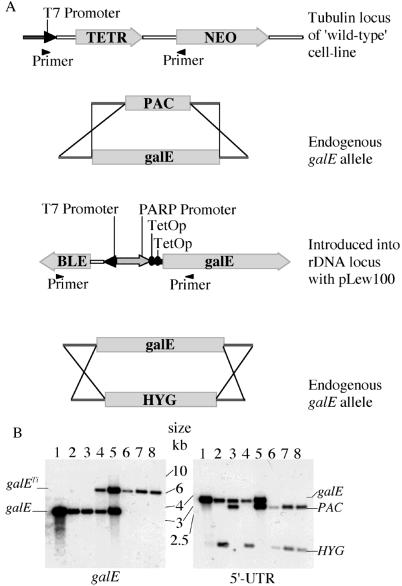
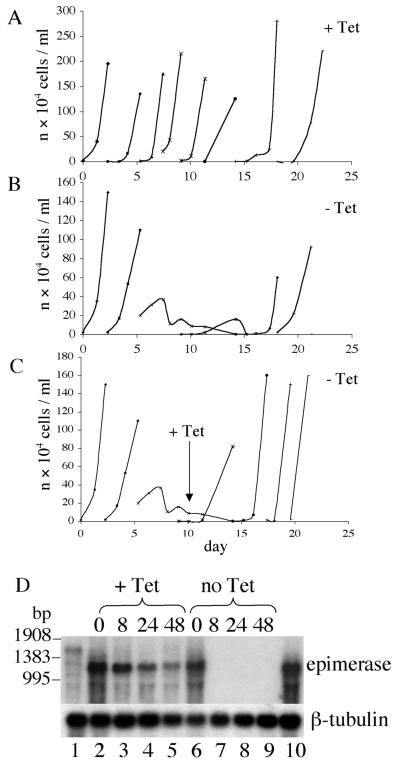
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
