

Phosphorylation and functional regulation of ClC-2 chloride channels expressed in Xenopus oocytes by M cyclin-dependent protein kinase
- PMID: 11986377
- PMCID: PMC2290285
- DOI: 10.1113/jphysiol.2001.016188
Phosphorylation and functional regulation of ClC-2 chloride channels expressed in Xenopus oocytes by M cyclin-dependent protein kinase
Abstract
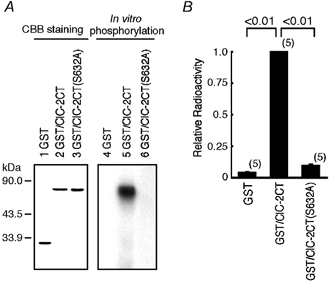
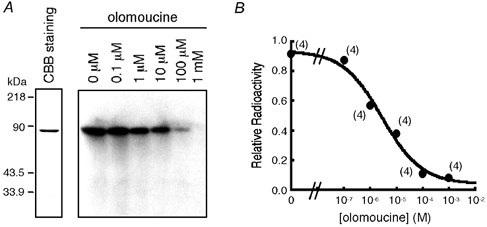
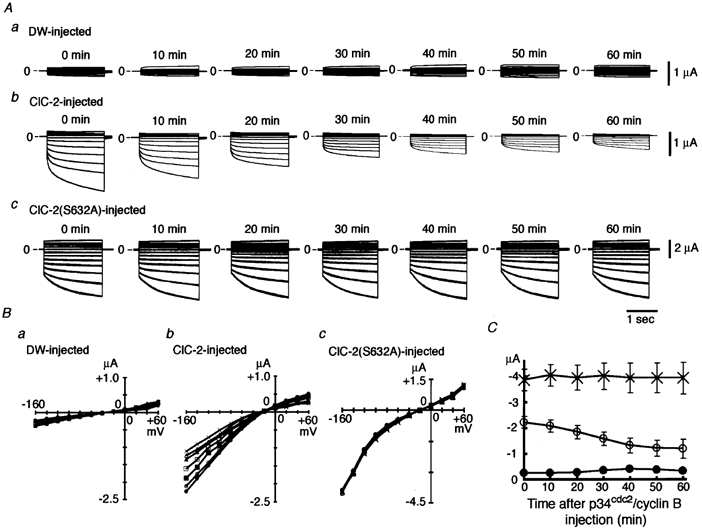
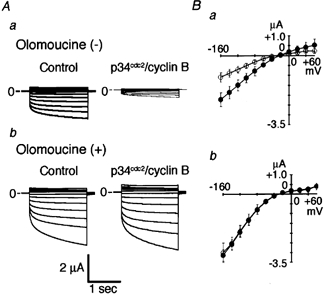
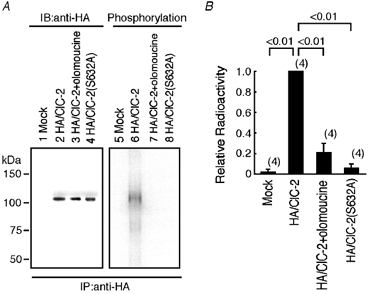
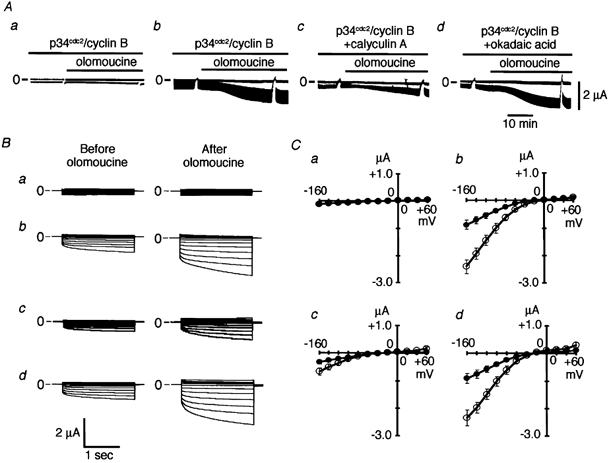
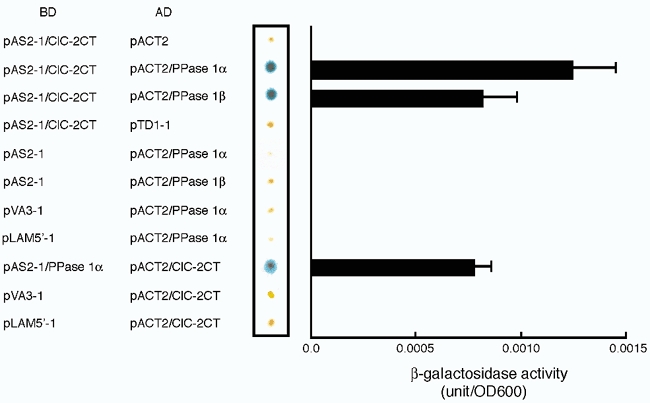
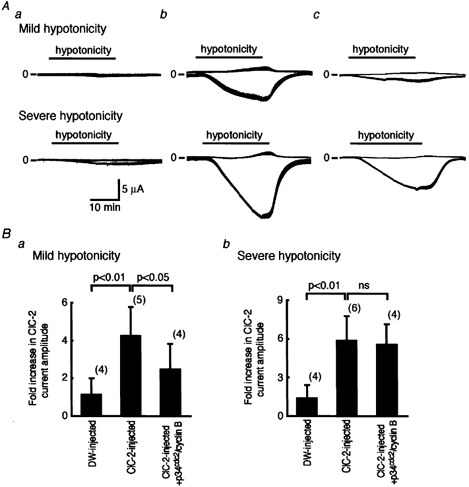
Many dramatic alterations in various cellular processes during the cell cycle are known to involve ion channels. In ascidian embryos and Caenorhabditis elegans oocytes, for example, the activity of inwardly rectifying Cl(-) channels is enhanced during the M phase of the cell cycle, but the mechanism underlying this change remains to be established. We show here that the volume-sensitive Cl(-) channel, ClC-2 is regulated by the M-phase-specific cyclin-dependent kinase, p34(cdc2)/cyclin B. ClC-2 channels were phosphorylated by p34(cdc2)/cyclin B in both in vitro and cell-free phosphorylation assays. ClC-2 phosphorylation was inhibited by olomoucine and abolished by a (632)Ser-to-Ala (S632A) mutation in the C-terminus, indicating that (632)Ser is a target of phosphorylation by p34(cdc2)/cyclin B. Injection of activated p34(cdc2)/cyclin B attenuated the ClC-2 currents but not the S632A mutant channel currents expressed in Xenopus oocytes. ClC-2 currents attenuated by p34(cdc2)/cyclin B were increased by application of the cyclin-dependent kinase inhibitor, olomoucine (100 microM), an effect that was inhibited by calyculin A (5 nM) but not by okadaic acid (5 nM). A yeast two-hybrid system revealed a direct interaction between the ClC-2 C-terminus and protein phosphatase 1. These data suggest that the ClC-2 channel is also counter-regulated by protein phosphatase 1. In addition, p34(cdc2)/cyclin B decreased the magnitude of ClC-2 channel activation caused by cell swelling. As the activities of both p34(cdc2)/cyclin B and protein phosphatase 1 vary during the cell cycle, as does cell volume, the ClC-2 channel could be regulated physiologically by these factors.
Figures
References
-
- Block ML, Moody WJ. A voltage-dependent chloride current linked to the cell cycle in ascidian embryos. Science. 1990;247:1090–1092. - PubMed
-
- Bubien JK, Kork KL, Rado TA, Frizzell RA. Cell cycle dependence of chloride permeability in normal and cystic fibrosis lymphocytes. Science. 1990;248:1416–1419. - PubMed
-
- Campbell KS, Buder A, Deuschle U. Interactions between the amino-terminal domain of p56lck and cytoplasmic domains of CD4 and CD8 alpha in yeast. European Journal of Immunology. 1995;25:2408–2412. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous