

Formation of a human immunodeficiency virus type 1 core of optimal stability is crucial for viral replication
- PMID: 11991995
- PMCID: PMC137032
- DOI: 10.1128/jvi.76.11.5667-5677.2002
Formation of a human immunodeficiency virus type 1 core of optimal stability is crucial for viral replication
Abstract
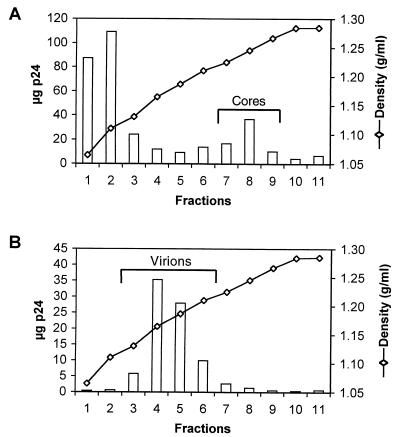
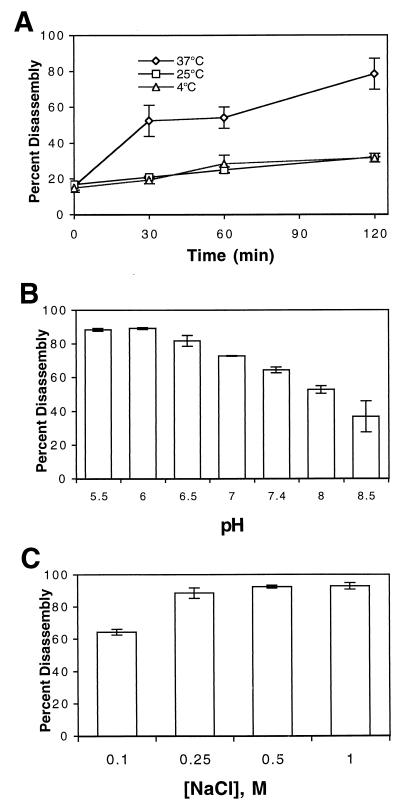
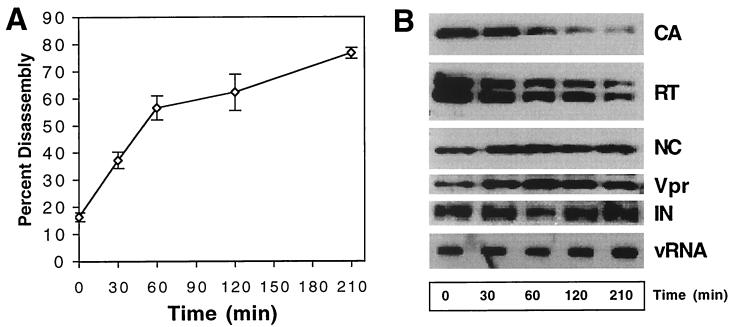
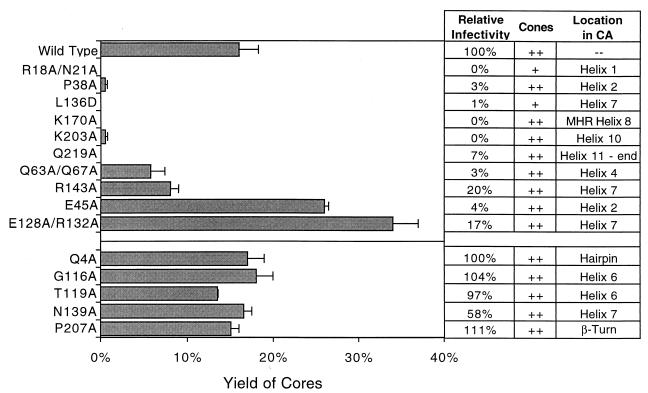
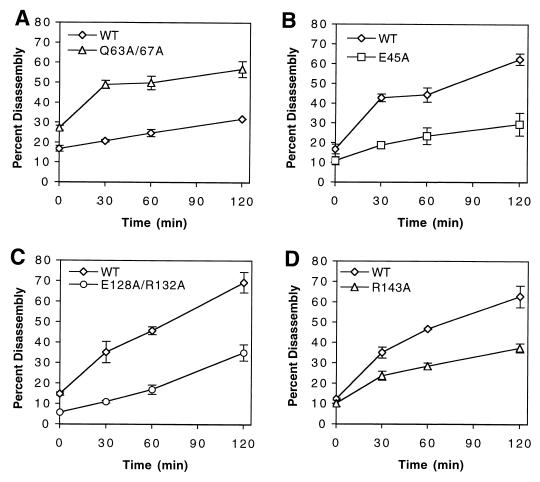
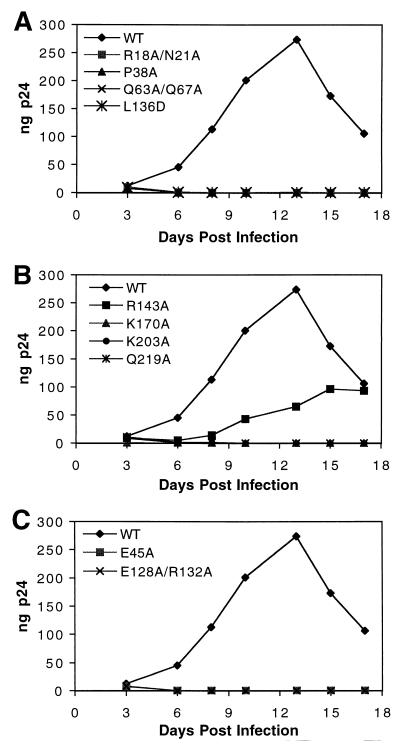
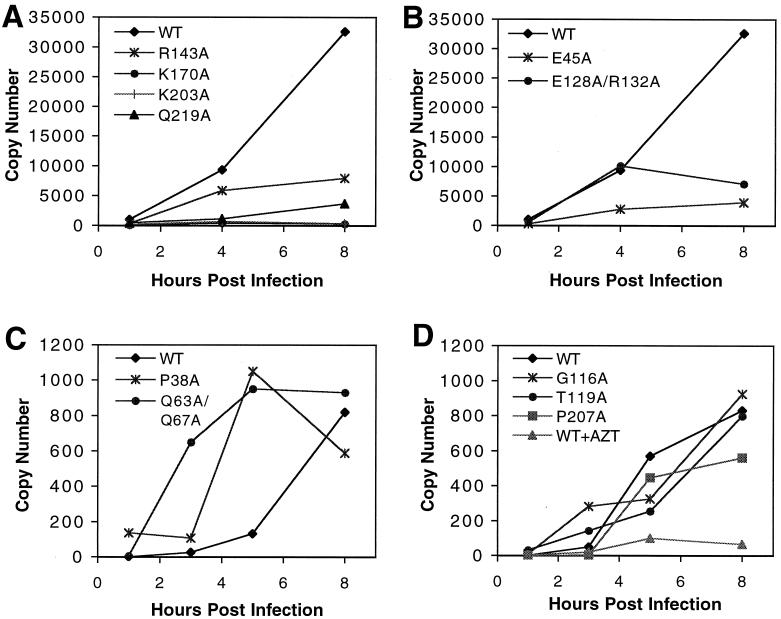
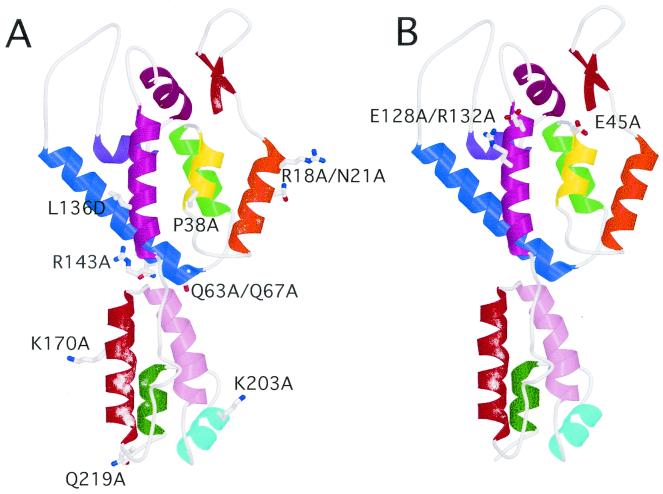
Virions of human immunodeficiency virus type 1 (HIV-1) and other lentiviruses contain conical cores consisting of a protein shell composed of the viral capsid protein (CA) surrounding an internal viral ribonucleoprotein complex. Although genetic studies have implicated CA in both early and late stages of the virus replication cycle, the mechanism of core disassembly following penetration of target cells remains undefined. Using quantitative assays for analyzing HIV-1 core stability in vitro, we identified point mutations in CA that either reduce or increase the stability of the HIV-1 core without impairing conical core formation in virions. Alterations in core stability resulted in severely attenuated HIV-1 replication and impaired reverse transcription in target cells with only minimal effects on viral DNA synthesis in permeabilized virions in vitro. We conclude that formation of a viral core of optimal stability is a prerequisite for efficient HIV-1 infection and suggest that disassembly of the HIV-1 core is a regulated step in infection that may be an attractive target for pharmacologic intervention.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
