

Polyploid measles virus with hexameric genome length
- PMID: 12006489
- PMCID: PMC126007
- DOI: 10.1093/emboj/21.10.2364
Polyploid measles virus with hexameric genome length
Abstract
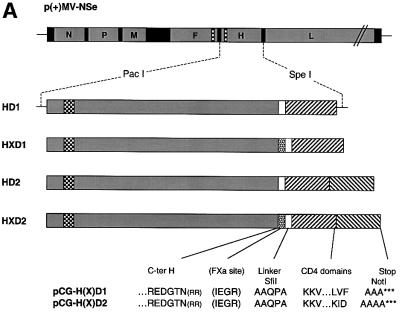
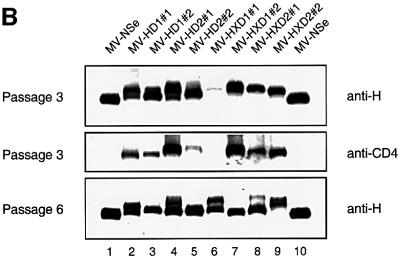
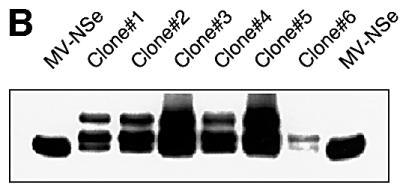
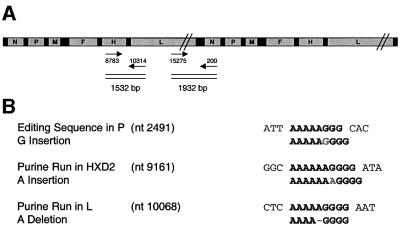
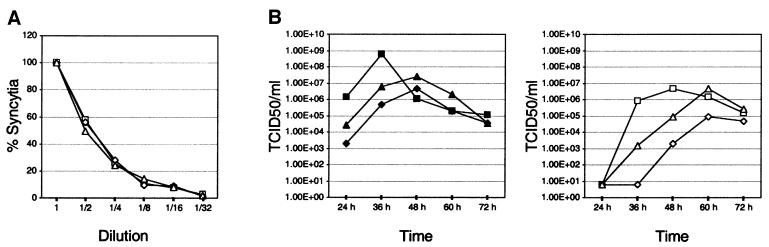
Particles of most virus species accurately package a single genome, but there are indications that the pleomorphic particles of parainfluenza viruses incorporate multiple genomes. We characterized a stable measles virus mutant that efficiently packages at least two genomes. The first genome is recombinant and codes for a defective attachment protein with an appended domain interfering with fusion-support function. The second has one adenosine insertion in a purine run that interrupts translation of the appended domain and restores function. In that genome, a one base deletion in a different purine run abolishes polymerase synthesis, but restores hexameric genome length, thus ensuring accurate RNA encapsidation, which is necessary for efficient replication. Thus, the two genomes are complementary. The infection kinetics of this mutant indicate that packaging of multiple genomes does not negatively affect growth. We also show that polyploid particles are produced in standard infections at no expense to infectivity. Our results illustrate how the particles of parainfluenza viruses efficiently accommodate cargoes of different volume, and suggest a mechanism by which segmented genomes may have evolved.
Figures

References
-
- Baczko K. et al. (1993) Clonal expansion of hypermutated measles virus in a SSPE brain. Virology, 197, 188–195. - PubMed
-
- Bluming A.Z. and Ziegler,J.L. (1971) Regression of Burkitt’s lymphoma in association with measles infection. Lancet, 2, 105–106. - PubMed
-
- Cathomen T., Buchholz,C.J., Spielhofer,P. and Cattaneo,R. (1995) Preferential initiation at the second AUG of the measles virus F mRNA: a role for the long untranslated region. Virology, 214, 628–632. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
