

Ceramide channels increase the permeability of the mitochondrial outer membrane to small proteins
- PMID: 12006562
- PMCID: PMC2246046
- DOI: 10.1074/jbc.M200754200
Ceramide channels increase the permeability of the mitochondrial outer membrane to small proteins
Abstract
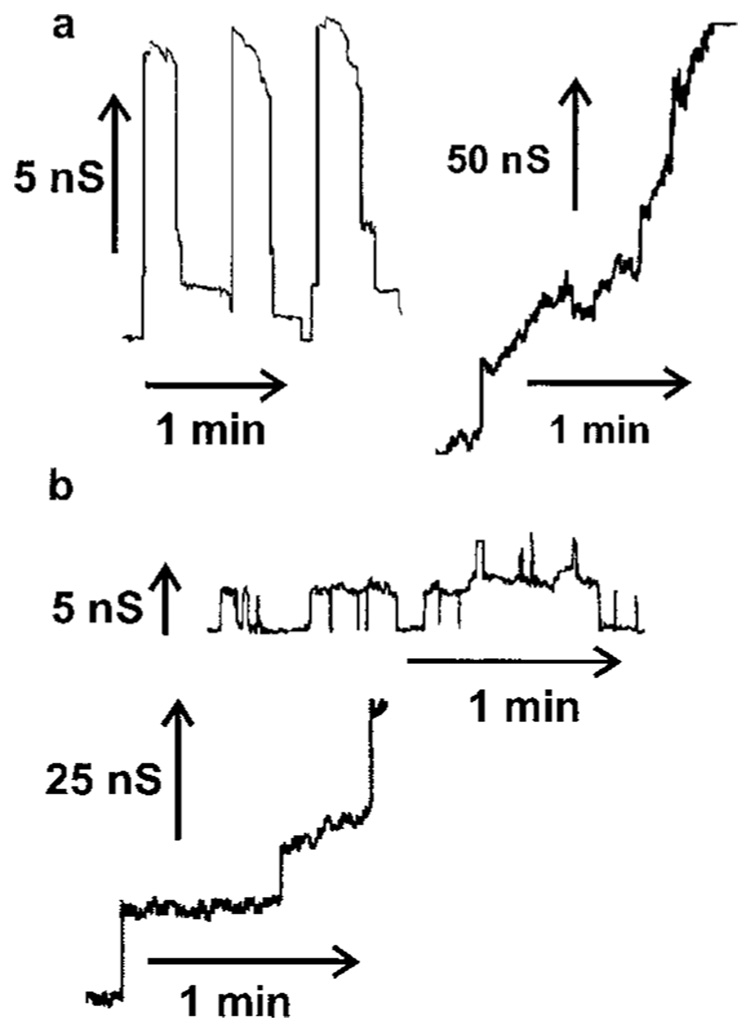
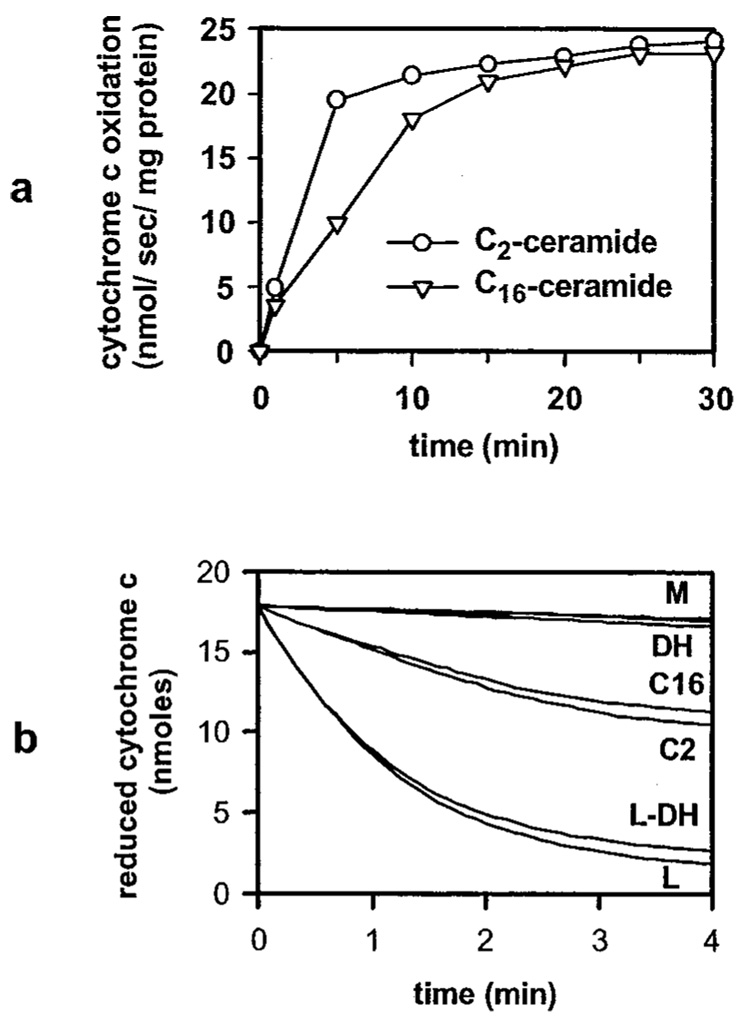
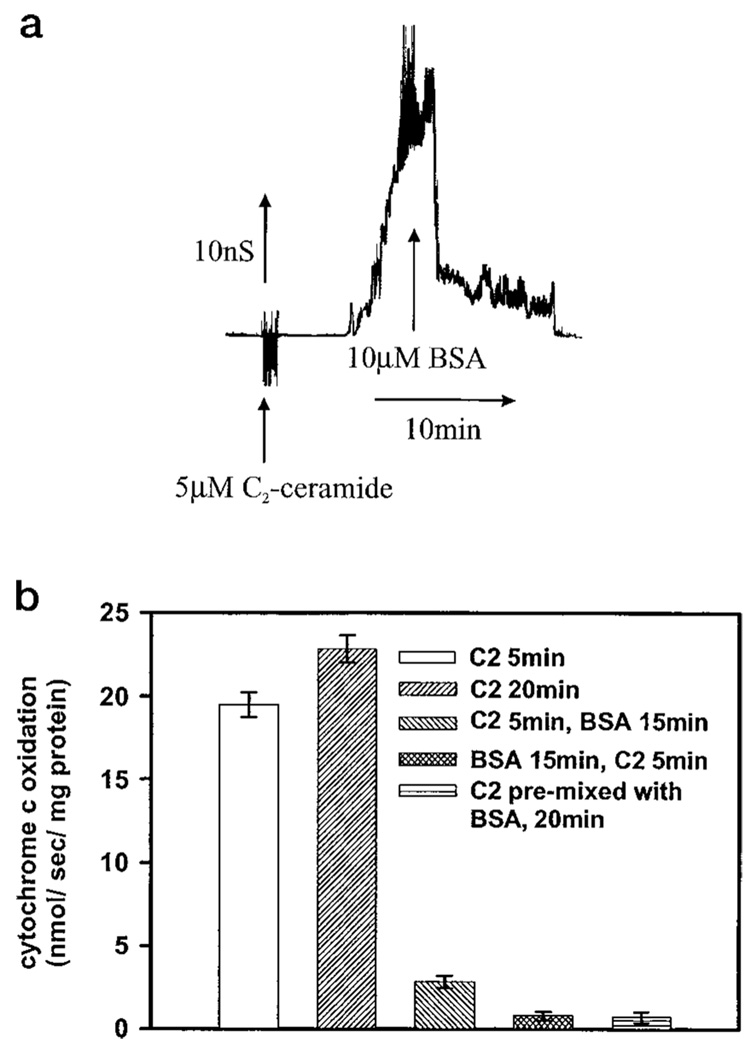
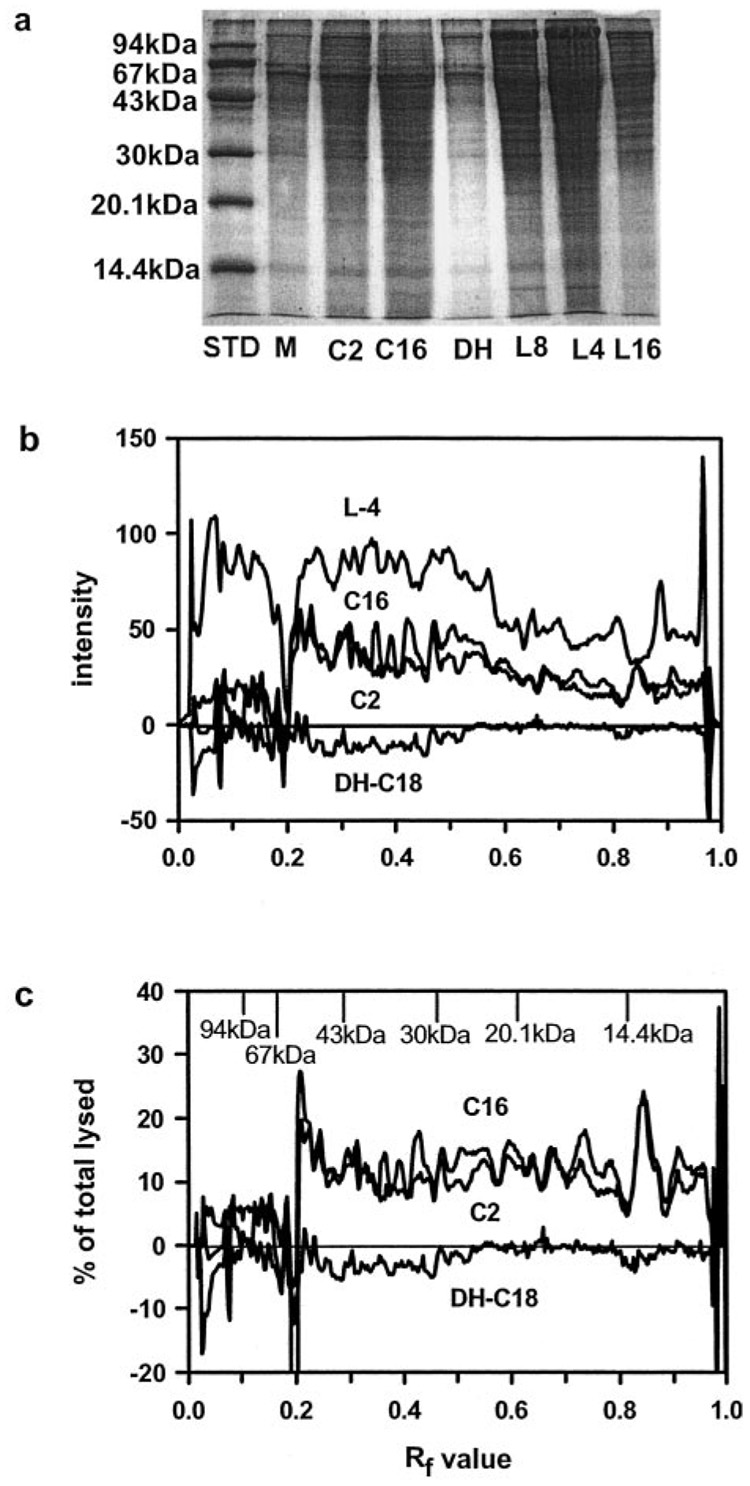
Ceramides are known to play a major regulatory role in apoptosis by inducing cytochrome c release from mitochondria. We have previously reported that C(2)- and C(16)-ceramide, but not dihydroceramide, form large channels in planar membranes (Siskind, L. J., and Colombini, M. (2001) J. Biol. Chem. 275, 38640-38644). Here we show that ceramides do not trigger a cytochrome c secretion or release mechanism, but simply raise the permeability of the mitochondrial outer membrane, via ceramide channel formation, to include small proteins. Exogenously added reduced cytochrome c was able to freely permeate the mitochondrial outer membrane with entry to and exit from the intermembrane space facilitated by ceramides in a dose- and time-dependent manner. The permeability pathways were eliminated upon removal of C(2)-ceramide by bovine serum albumin, thus ruling out a detergent-like effect of C(2)-ceramide on membranes. Ceramide channels were not specific to cytochrome c, as ceramides induced release of adenylate kinase, but not fumerase from isolated mitochondria, showing some specificity of these channels for the outer mitochondrial membrane. SDS-PAGE results show that ceramides allow release of intermembrane space proteins with a molecular weight cut-off of about 60,000. These results indicate that the ceramide-induced membrane permeability increases in isolated mitochondria are via ceramide channel formation and not a release mechanism, as the channels that allow cytochrome c to freely permeate are reversible, and are not specific to cytochrome c.
Figures
References
-
- Radin NS. Eur. J. Biochem. 2001;268:193–204. - PubMed
-
- Kolesnick RN, Krönke M. Annu. Rev. Physiol. 1998;60:643–665. - PubMed
-
- Ariga T, Jarvis WD, Yu RK. J. Lipid Res. 1998;39:1–16. - PubMed
-
- Hannun YA. Science. 1996;274:1855–1859. - PubMed
-
- Kolesnick RN, Goñi FM, Alonoso A. J. Cell. Physiol. 2000;184:285–300. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
