

Review
doi: 10.1128/IAI.70.6.2741-2751.2002.
Immunity to murine chlamydial genital infection
Affiliations
- PMID: 12010958
- PMCID: PMC128027
- DOI: 10.1128/IAI.70.6.2741-2751.2002
Item in Clipboard
Review
Immunity to murine chlamydial genital infection
Infect Immun.
2002 Jun.
No abstract available
Figures
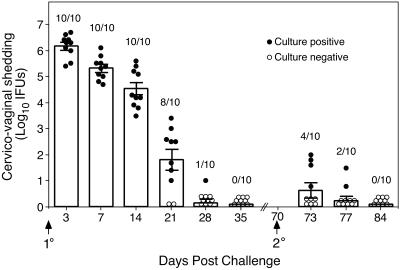
C. trachomatis MoPn genital tract infection of female C57BL/6 mice. Mice were infected intravaginally with 100 50% infectious doses (approximately 103 inclusion-forming units [IFUs]) of MoPn EBs at day 0, and IFUs were enumerated from swabs of the vaginal vault at the indicated times post primary and secondary infectious challenge. Data are presented as the mean numbers of IFUs recovered from all mice at each time point (indicated by the bar graph) and the standard errors of the means. Circles, the number of IFUs recovered from individual mice; closed circles, culture-positive mice; open circles, culture-negative animals. Values above the bars indicate the numbers of culture-positive animals per total number infected at each time point post-primary and -secondary challenge. Following the resolution of primary infection only 4 of 10 (40%) mice were infectable upon secondary challenge, thus demonstrating that solid immunity develops in the vast majority of mice. Furthermore, mice that were reinfected showed marked diminution of bacterial load and a vastly shortened course of infection. The duration of infection was 7 days or less (compared to 4 to 5 weeks for primary infection) and there was a 5.5 log10 reduction in recoverable IFUs in culture-positive reinfected mice (3 days post-secondary challenge compared to 3 days post-primary challenge).
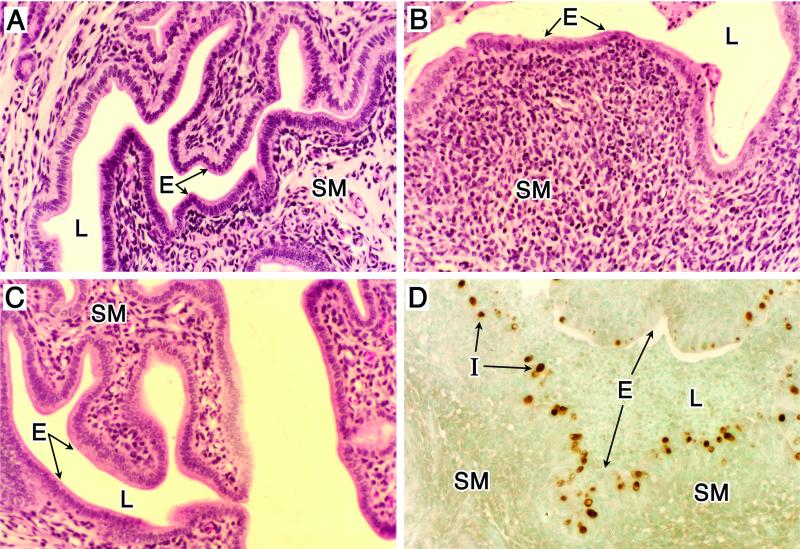
Histopathology of genital tract tissue of C57BL/6 female mice following infection with C. trachomatis (MoPn). Animals were infected as described in the legend for Fig. 1. (A to C) Uterine tissues stained with hematoxylin and eosin. (A) Noninfected. (B) Day 14 postinfection. (C) Day 42 postinfection. Note the marked thinning of the epithelial surface accompanied by an intense subacute inflammatory response consisting of polymorphonuclear cells, macrophages, and lymphocytes (B). The inflammatory response subsides as infection resolves (C). Localization of chlamydial inclusions to the epithelium was demonstrated by staining uterine tissue, obtained from mice at day 7 postinfection, with anti-MoPn MOMP immunoperoxidase (D). E, epithelium; L, lumen; I, chlamydial inclusion; SM, submucosae. Magnifications, ×100. This figure was adapted with permission from reference .
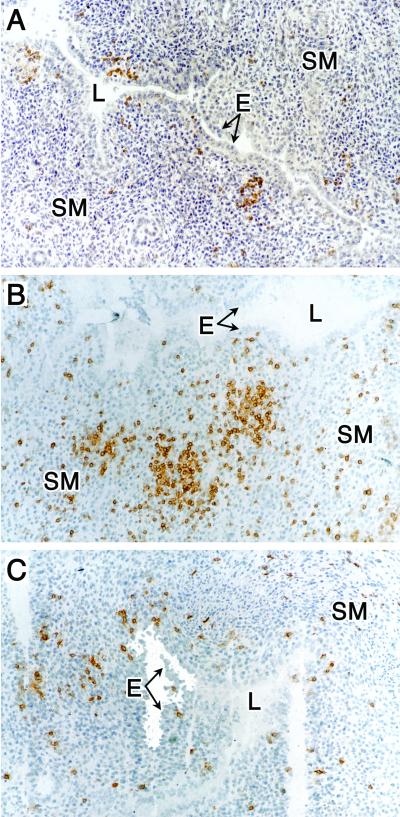
Immunohistochemical staining of B cells and T-cell subpopulations in uterine tissue from chlamydiae-infected mice at 14 days postinfection. (A) Anti-CD45R (B cells). (B) Anti-CD4 (CD4+ T cells). (C) Anti-CD8 (CD8+ T cells). Magnification, ×100. See the legend to Fig. 2 for abbreviations. This figure was adapted with permission from reference .
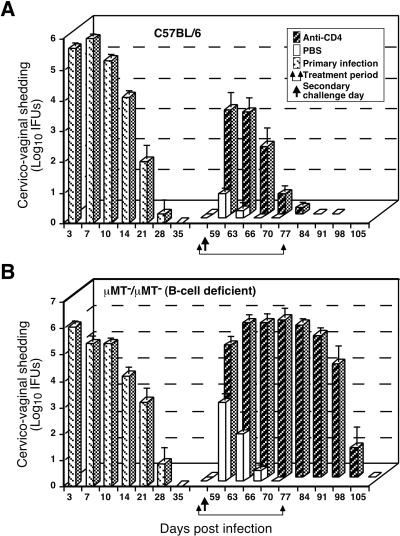
Effect of anti-CD4 treatment on the resolution of secondary C. trachomatis MoPn genital tract infection of wild-type C57BL/6 mice (A) and μMT/μMT B-cell-deficient gene knockout mice (B). Mice were infected and IFUs were enumerated as described in the legend to Fig. 1. Following the resolution of primary infection, immune mice were either treated with phosphate-buffered saline (PBS) or anti-CD4 monoclonal antibody and then rechallenged. Mice depleted of CD4+ T cells resolve secondary infection, although resolution is delayed (A), whereas mice depleted of both CD4+ T cells and B cells fail to resolve secondary infection. These data convincingly demonstrate a role for B cells and CD4+ T cells in adaptive immunity to chlamydial genital tract infection. This figure was adapted with permission from reference .
References
-
- Allan, I., and J. H. Pearce. 1983. Amino acid requirements of strains of Chlamydia trachomatis and C. psittaci growing in McCoy cells: relationship with clinical syndrome and host origin. J. Gen. Microbiol. 129:2001-2007. - PubMed
-
- Anttila, T., P. Saikku, P. Koskela, A. Bloigu, J. Dillner, I. Ikaheimo, E. Jellum, M. Lehtinnen, P. Lenner, T. Hakulinen, A. Narvanen, E. Pukkala, S. Thoresen, L. Youngman, and J. Paavonen. 2001. Serotypes of Chlamydia trachomatis and risk for development of cervical squamous cell carcinoma. JAMA 285:47-51. - PubMed
-
- Barron, A. L., H. J. White, R. G. Rank, B. L. Soloff, and E. B. Moses. 1981. A new animal model for the study of Chlamydia trachomatis genital infections: infection of mice with the agent of mouse pneumonitis. J. Infect. Dis. 143: 63-66. - PubMed
-
- Beatty, P. R., and R. S. Stephens. 1994. CD8+ T lymphocyte-mediated lysis of Chlamydia-infected L cells using an endogenous antigen pathway. J. Immunol. 153:4588-4595. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
