

Characterization of FRO1, a pea ferric-chelate reductase involved in root iron acquisition
- PMID: 12011340
- PMCID: PMC155873
- DOI: 10.1104/pp.010829
Characterization of FRO1, a pea ferric-chelate reductase involved in root iron acquisition
Abstract
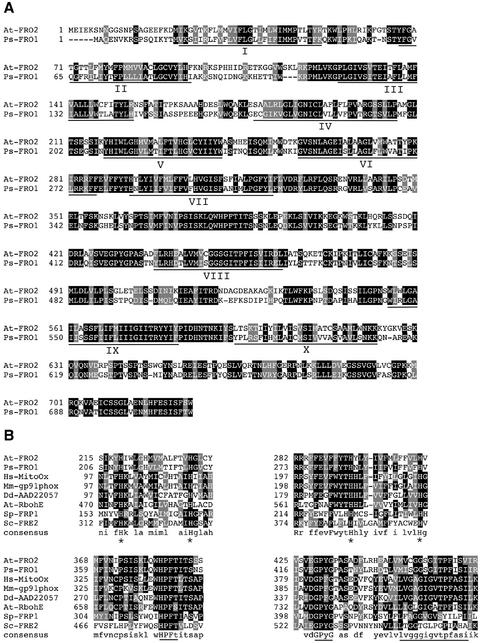
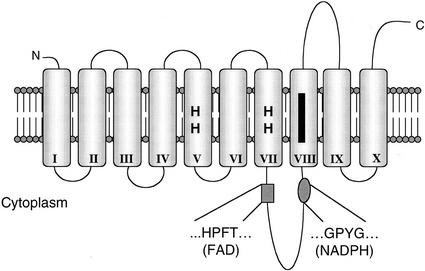
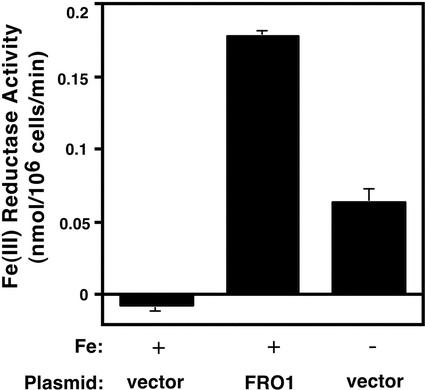
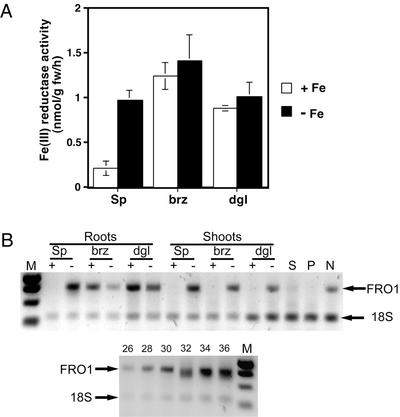
To acquire iron, many plant species reduce soil Fe(III) to Fe(II) by Fe(III)-chelate reductases embedded in the plasma membrane of root epidermal cells. The reduced product is then taken up by Fe(II) transporter proteins. These activities are induced under Fe deficiency. We describe here the FRO1 gene from pea (Pisum sativum), which encodes an Fe(III)-chelate reductase. Consistent with this proposed role, FRO1 shows similarity to other oxidoreductase proteins, and expression of FRO1 in yeast conferred increased Fe(III)-chelate reductase activity. Furthermore, FRO1 mRNA levels in plants correlated with Fe(III)-chelate reductase activity. Sites of FRO1 expression in roots, leaves, and nodules were determined. FRO1 mRNA was detected throughout the root, but was most abundant in the outer epidermal cells. Expression was detected in mesophyll cells in leaves. In root nodules, mRNA was detected in the infection zone and nitrogen-fixing region. These results indicate that FRO1 acts in root Fe uptake and they suggest a role in Fe distribution throughout the plant. Characterization of FRO1 has also provided new insights into the regulation of Fe uptake. FRO1 expression and reductase activity was detected only in Fe-deficient roots of Sparkle, whereas both were constitutive in brz and dgl, two mutants with incorrectly regulated Fe accumulation. In contrast, FRO1 expression was responsive to Fe status in shoots of all three plant lines. These results indicate differential regulation of FRO1 in roots and shoots, and improper FRO1 regulation in response to a shoot-derived signal of iron status in the roots of the brz and dgl mutants.
Figures
References
-
- Appel RD, Bairoch A, Hochstrasser DF. A new generation of information retrieval tools for biologists: the example of the ExPASy WWW server. Trends Biochem Sci. 1994;19:258–260. - PubMed
-
- Brown JC, Chaney RL, Amber JE. A new tomato mutant inefficient in the transport of iron. Physiol Plant. 1971;25:45–53.
-
- Bughio N, Takahashi M, Yoshimura E, Nishizawa NK, Mori S. Characteristics of light-regulated iron transport system in barley chloroplasts. Soil Sci Plant Nutr. 1997;43:959–963.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
