

The biophysics of leaf growth in salt-stressed barley. A study at the cell level
- PMID: 12011367
- PMCID: PMC155900
- DOI: 10.1104/pp.001164
The biophysics of leaf growth in salt-stressed barley. A study at the cell level
Abstract
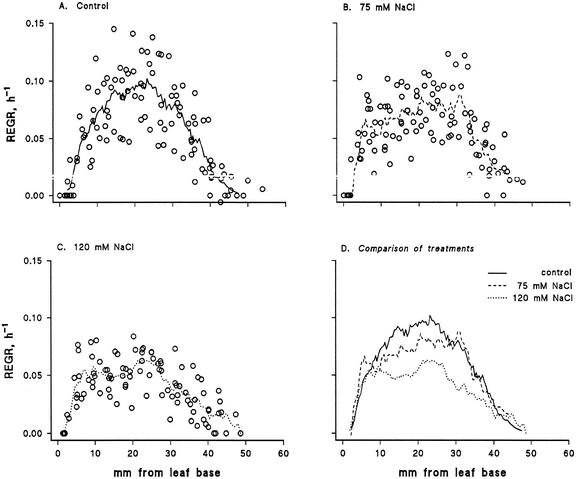
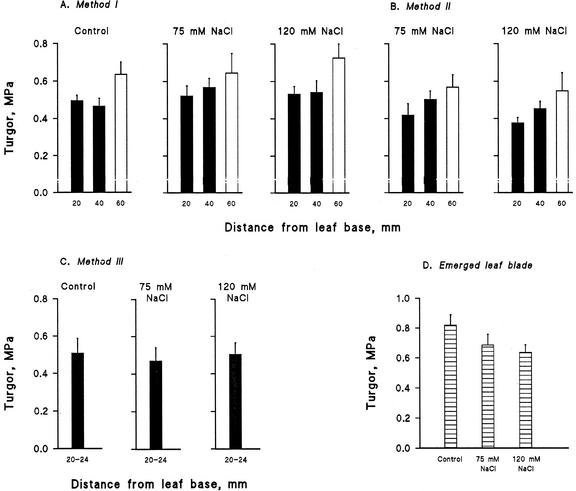
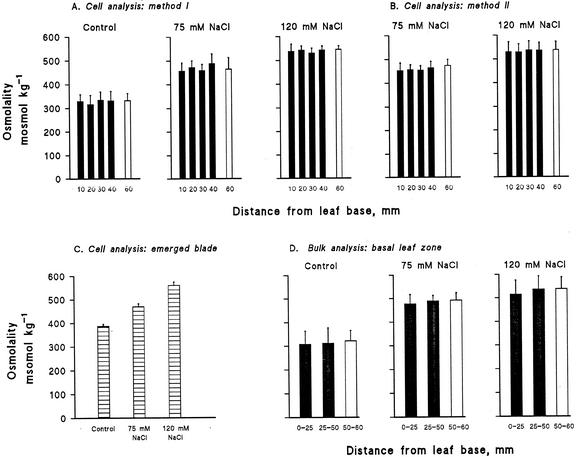
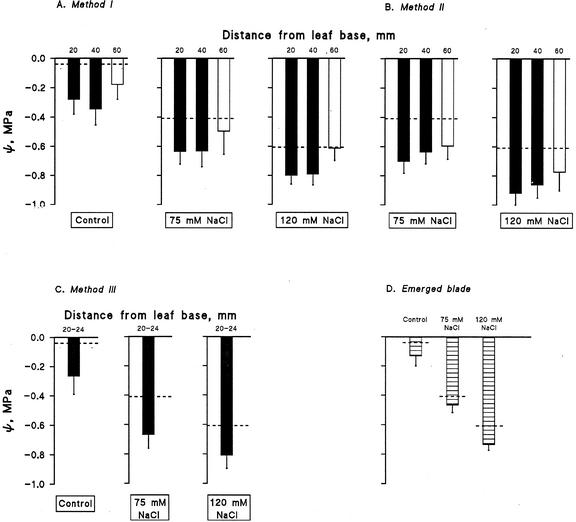
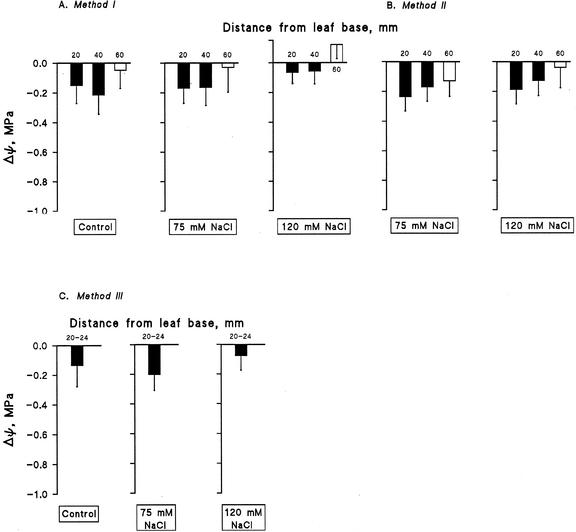
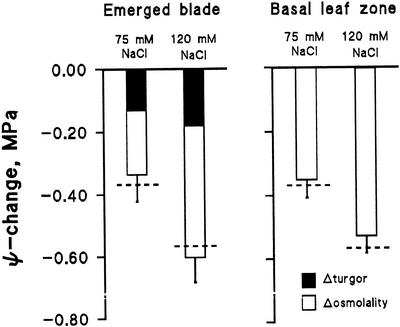
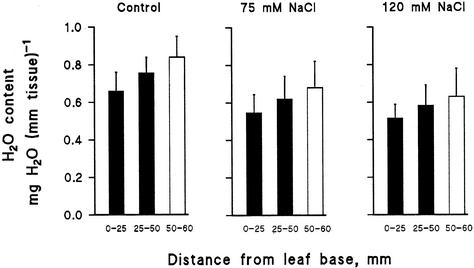
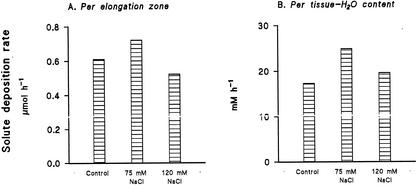
Biophysical parameters potentially involved in growth regulation were studied at the single-cell level in the third leaf of barley (Hordeum vulgare) after exposure to various degrees of NaCl stress for 3 to 5 d. Gradients of elongation growth were measured, and turgor pressure, osmolality, and water potentials (psi) were determined (pressure probe and picoliter osmometry) in epidermal cells of the elongation zone and the mature blade. Cells in the elongation zone adjusted to decreasing external psi through increases in cell osmolality that were accomplished by increased solute loads and reduced water contents. Cell turgor changed only slightly. In contrast, decreases in turgor also contributed significantly to psi adjustment in the mature blade. Solute deposition rates in the elongation zone increased at moderate stress levels as compared with control conditions, but decreased again at more severe NaCl exposure. Growth-associated psi gradients between expanding epidermal cells and the xylem were significant under control and moderate stress conditions (75 mM NaCl) but seemed negligible at severe stress (120 mM NaCl). We conclude that leaf cell elongation in NaCl-treated barley is probably limited by the rate at which solutes can be taken up to generate turgor, particularly at high NaCl levels.
Figures
References
-
- Arif H, Tomos AD. Control of wheat leaf growth under saline conditions. In: Lieth H, Al Masoom A, editors. Towards the Rational Use of High Salinity Tolerant Plants. Vol. 2. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1993. pp. 45–52.
-
- Barlow EWR. Water relations of expanding leaves. Aust J Plant Physiol. 1986;13:45–58.
-
- Boyer JS. Water transport in plants: mechanisms of apparent changes in resistance during absorption. Planta. 1974;117:187–207. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
