

ARGONAUTE1 is required for efficient RNA interference in Drosophila embryos
- PMID: 12011447
- PMCID: PMC124499
- DOI: 10.1073/pnas.072190799
ARGONAUTE1 is required for efficient RNA interference in Drosophila embryos
Abstract
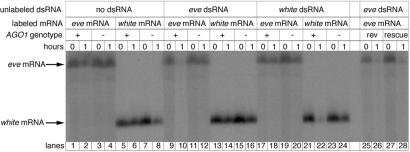
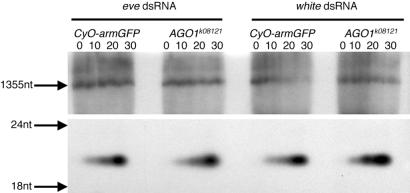
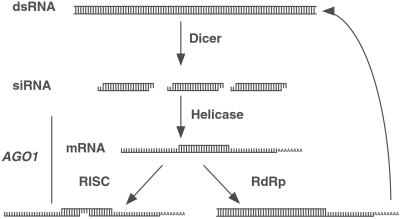
Double-stranded RNA (dsRNA) triggers homology-dependent posttranscriptional gene interference (RNAi) in a diverse range of eukaryotic organisms, in a process mechanistically related to viral and transgene-mediated cosuppression. RNAi is characterized by the conversion of long dsRNA into approximately 21-25-nt small interfering RNAs (siRNA) that guide the degradation of homologous mRNA. Many of the genes required for siRNA production and target mRNA degradation are widely conserved. Notably, members of the Argonaute-like gene family from Arabidopsis, Caenorhabditis elegans, Drosophila, and Neurospora have been genetically and/or biochemically identified as components of the RNAi/cosuppression pathway. We show here that mutations in the Drosophila Argonaute1 (AGO1) gene suppress RNAi in embryos. This defect corresponds to a reduced ability to degrade mRNA in response to dsRNA in vitro. Furthermore, AGO1 is not required for siRNA production in vitro nor can the introduction of siRNA bypass AGO1 mutants in vivo. These data suggest that AGO1 functions downstream of siRNA production.
Figures
References
-
- Hammond S M, Caudy A A, Hannon G J. Nat Rev Genet. 2001;2:110–119. - PubMed
-
- Fire A, Xu S, Montgomery M K, Kostas S A, Driver S E, Mello C C. Nature (London) 1998;391:806–811. - PubMed
-
- Grishok A, Pasquinelli A E, Conte D, Li N, Parrish S, Ha I, Baillie D L, Fire A, Ruvkun G, Mello C C. Cell. 2001;106:23–34. - PubMed
-
- Hutvagner G, McLachlan J, Pasquinelli A E, Balint E, Tuschl T, Zamore P D. Science. 2001;293:834–838. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
