

Conformation-dependent regulation of inward rectifier chloride channel gating by extracellular protons
- PMID: 12015423
- PMCID: PMC2290315
- DOI: 10.1113/jphysiol.2002.016485
Conformation-dependent regulation of inward rectifier chloride channel gating by extracellular protons
Abstract
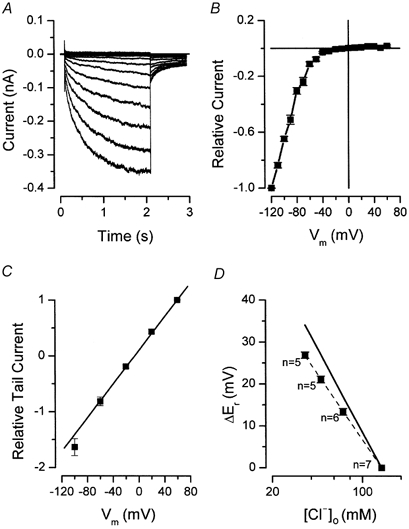
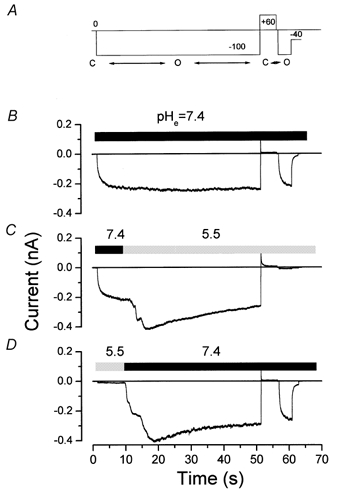
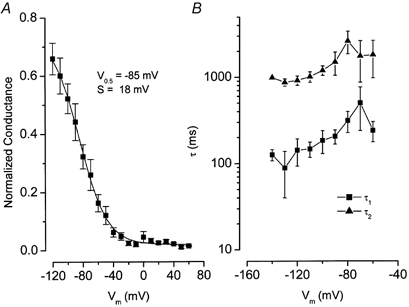
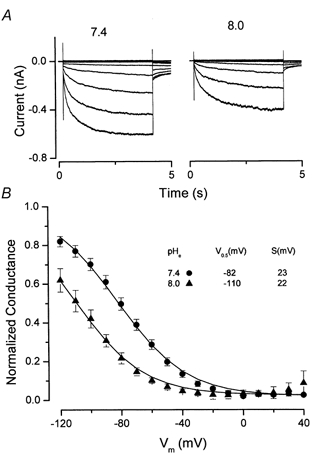
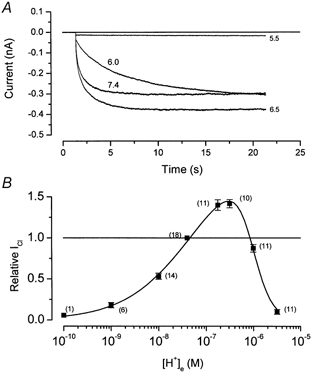
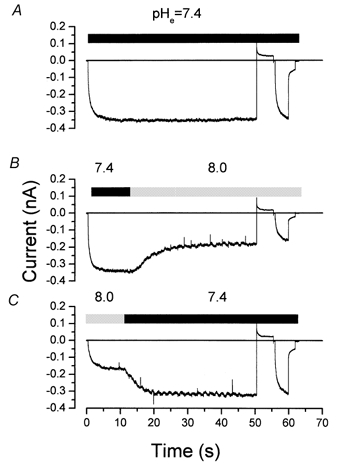
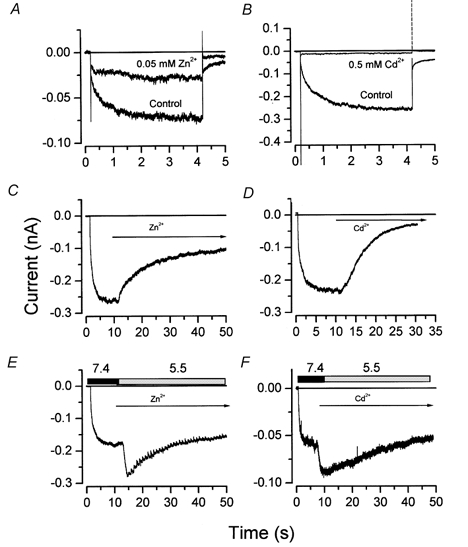
We have investigated the gating properties of the inward rectifier chloride channel (Cl(ir)) from mouse parotid acinar cells by external protons (H(+)(o)) using the whole-cell patch-clamp technique. Increasing the pH(o) from 7.4 to 8.0 decreased the magnitude of Cl(ir) current by shifting the open probability to more negative membrane potentials with little modification of the activation kinetics. The action of elevated pH was independent of the conformational state of the channel. The effects of low pH on Cl(ir) channels were dependent upon the conformational state of the channel. That is, application of pH 5.5 to closed channels essentially prevented channel opening. In contrast, application of pH 5.5 to open channels actually increased the current. These results are consistent with the existence of two independent protonatable sites: (1) a site with a pK near 7.3, the titration of which shifts the voltage dependence of channel gating; and (2) a site with pK = 6.0. External H(+) binds to this latter site (with a stoichiometry of two) only when the channels are closed and prevent channel opening. Finally, block of channels by Zn(2+) and Cd(2+) was inhibited by low pH media. We propose that mouse parotid Cl(ir) current has a bimodal dependence on the extracellular proton concentration with maximum activity near pH 6.5: high pH decreases channel current by shifting the open probability to more negative membrane potentials and low pH also decreases the current but through a proton-dependent stabilization of the channel closed state.
Figures
References
-
- Carew MA, Thorn P. Identification of ClC-2-like chloride currents in pig pancreatic acinar cells. Pflügers Archiv. 1996;433:84–90. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources