

Review
doi: 10.1038/417343a.
A small-systems approach to motor pattern generation
Affiliations
- PMID: 12015615
- PMCID: PMC6494453
- DOI: 10.1038/417343a
Item in Clipboard
Review
A small-systems approach to motor pattern generation
Nature.
.
Abstract
How neuronal networks enable animals, humans included, to make coordinated movements is a continuing goal of neuroscience research. The stomatogastric nervous system of decapod crustaceans, which contains a set of distinct but interacting motor circuits, has contributed significantly to the general principles guiding our present understanding of how rhythmic motor circuits operate at the cellular level. This results from a detailed documentation of the circuit dynamics underlying motor pattern generation in this system as well as its modulation by individual transmitters and neurons.
Figures
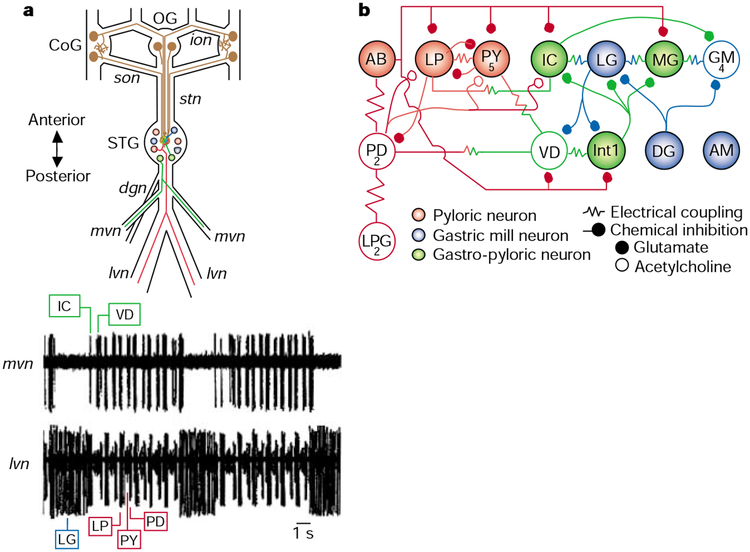
The gastric mill and pyloric circuits in the stomatogastric nervous system. a, Schematic of the stomatogastric nervous system in the crab Cancer borealis (top panel). The neurons with somata in the commissural (CoG) and oesophageal (OG) ganglia represent modulatory projection neurons that influence the gastric mill (blue), pyloric (red) and gastro-pyloric (green) neurons in the stomatogastric ganglion (STG). Gastro-pyloric neurons are active with both the gastric mill and the pyloric rhythms. Most STG neurons are motor neurons that project axons posteriorly through the lateral (lvn) or medial (mvn) ventricular nerves. Illustrated are two STG neurons that project through these nerves. The bottom panel shows that the gastric mill and pyloric rhythms are readily recorded extracellularly via these two nerves in situ (recordings modified from ref. 12). Comparable recordings are routinely made in vitro (in completely isolated stomatogastric nervous system). The gastric mill rhythm is represented by the relatively long duration, rhythmic action potential bursts in the lateral gastric (LG) neuron and the concomitant elimination of activity in the inferior cardiac (IC) and ventricular dilator (VD) neurons. The faster pyloric rhythm is evident from the sequentially repeating action potential bursts in the lateral pyloric (LP), pyloric (PY), pyloric dilator (PD), IC and VD neurons. The activity in the LP, PY and PD neurons is obscured during each LG neuron burst. Additional abbreviations: dgn, dorsal gastric nerve; ion, inferior oesophageal nerve; son, superior oesophageal nerve; stn, stomatogastric nerve. b, Schematic of the STG neural network the crab Cancer borealis, showing gastric mill and pyloric circuits. The pyloric neurons (red) are shown in their normal sequence of pyloric-timed activity, with time progressing from left to right. The neurons that show gastric mill-timed activity are represented so that the neurons active during protraction of the teeth are on top, and those active during retraction of the teeth are below. Neurons labelled as ‘gastro-pyloric neurons’ exhibit both gastric mill- and pyloric-timed activity. Abbreviations: AB, anterior burster neuron; MG, medial gastric neuron; GM, gastric mill neuron; Int1, Interneuron 1; DG, dorsal gastric neuron. Adapted from ref. .
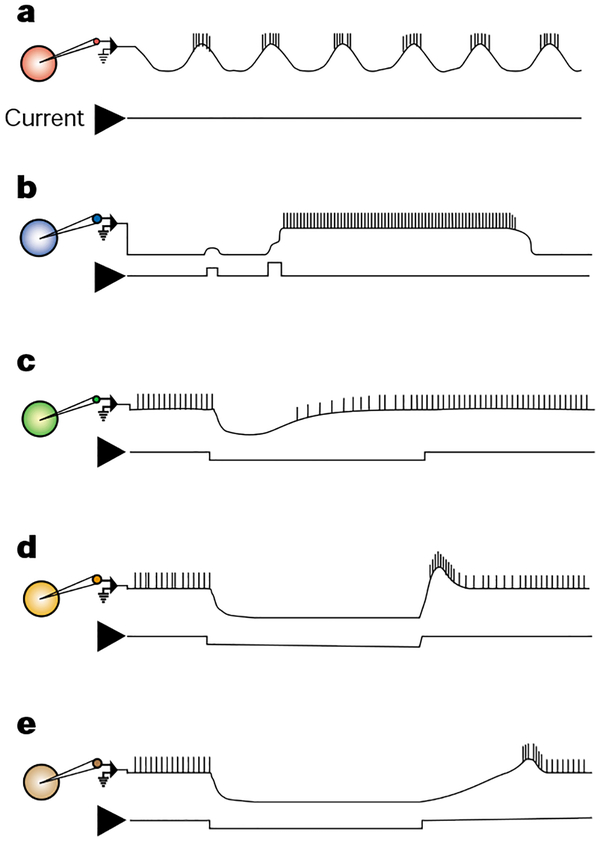
Illustrations of some of the membrane properties occurring in intracellularly recorded neurons (coloured circles) of central pattern generating circuits, including those in the STG. Each current trace is a monitor of depolarizing (up) and/or hyperpolarizing (down) current injections into the recorded neuron. Comparable events can be elicited by excitatory and inhibitory synaptic input. a, An endogenous oscillator undergoes rhythmic membrane potential oscillations without requiring any rhythmic input. b, The plateau potential is a persistent depolarizing response that generates persistent action potential activity and outlasts the triggering stimulus. It has a membrane potential threshold for its activation and it eventually self-terminates. c, Escape from inhibition is an excitation that is triggered by a sufficient amplitude and duration of hyperpolarization. It enables the neuron to depolarize, often to the point of firing action potentials, despite the continued presence of a hyperpolarizing input. d, Post-inhibitory rebound (PIR) is also triggered by hyperpolarization, but it represents an overshooting of the resting potential with an associated burst of action potentials after termination of the hyperpolarization. e, PIR delay shows a slower rebound depolarization, compared with PIR, after a period of hyperpolarization.
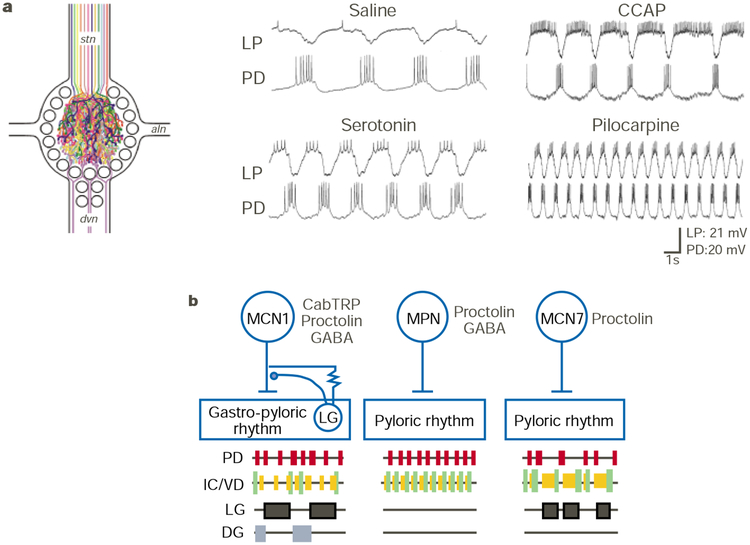
Modulation of the pyloric and gastric mill rhythms in the STG. a, Schematic of the modulatory innervation of the STG (left panel; adapted from ref. 4). Each colour represents a distinct neuromodulatory transmitter or complement of transmitters localized to the axon and STG terminals of projection neurons that innervate the STG via the stomatogastric nerve (stn) and of sensory neurons that innervate the STG via the dorsal ventricular nerve (dvn). Individual bath application of different modulators to the isolated STG in the crab Cancer borealis elicits different versions of the pyloric rhythm (right panel; S. R. Hertzberg, M.P.B. & M.P.N., unpublished data). The pyloric rhythm is monitored by intracellular recordings of two pyloric circuit neurons (PD and LP). Modulator concentrations: serotonin (10−5 M), crustacean cardioactive peptide (CCAP; 10−6 M), and the muscarinic agonist pilocarpine (10−5 M). Each application was followed by a saline wash, during which the rhythm returned to control conditions (saline). All recordings are from the same preparation. b, Selective activation of distinct modulatory projection neurons which have a peptide co-transmitter in common (proctolin) have different actions on the pyloric and gastric mill circuits in the STG. Note that the inhibitory and electrical synapses from the LG neuron to MCN1 occur within the STG neuropil,. Abbreviations: CabTRP, Cancer borealis tachykinin-related peptide; MCN1/7, modulatory commissural neuron 1/7; MPN, modulatory proctolin neuron. Modified from ref. .
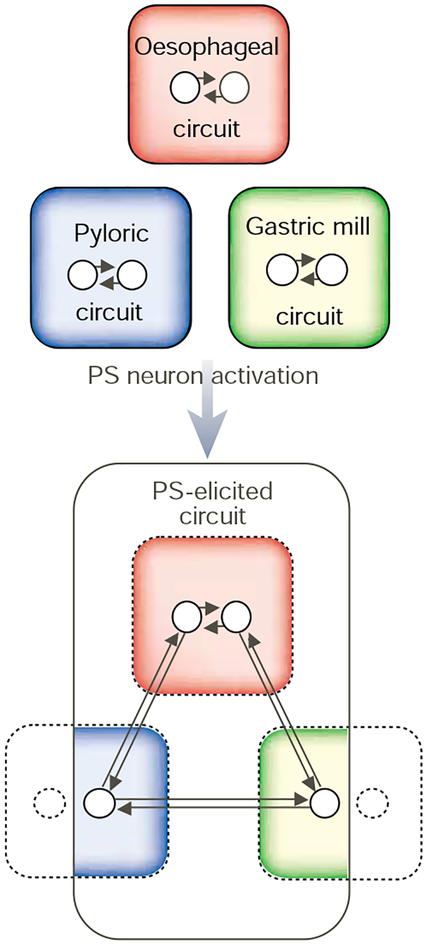
Modulatory input can dismantle and reconfigure the STG circuits. Generally, the gastric mill, pyloric and oesophageal circuits generate distinct motor rhythms. However, activation of some modulatory inputs, such as the pyloric suppressor (PS) neuron, can eliminate these rhythms and replace them with a single, conjoint motor pattern. The PS-elicited motor circuit includes a subset of the neurons (circles within coloured regions) that comprise the three circuits. The PS neuron also inhibits the activity of the gastric mill and pyloric circuit neurons (circles with dashed outlines located outside coloured regions) that do not participate in this conjoint motor pattern. Adapted from ref. ; see also ref. .
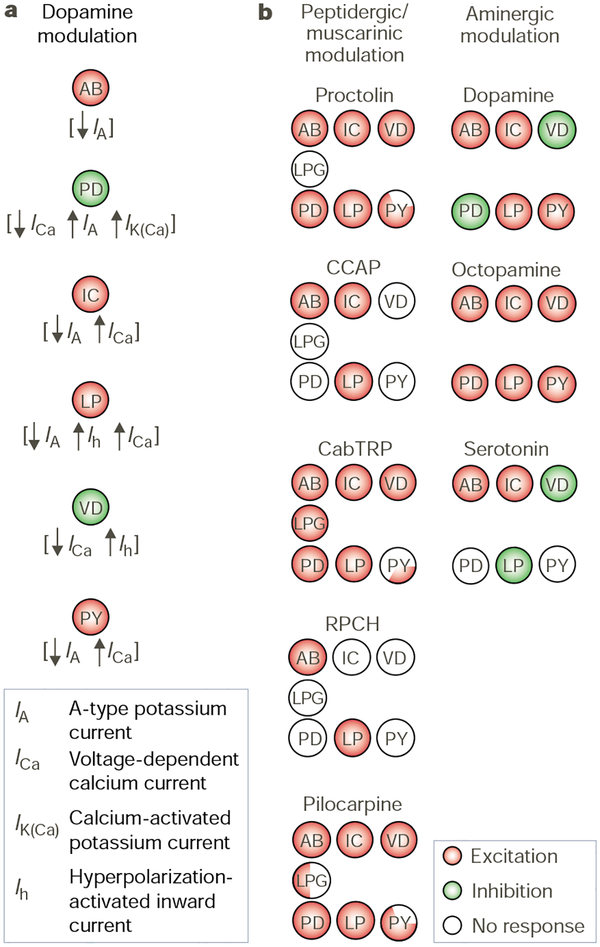
Convergence and divergence of transmitter actions. a, Dopamine influences a distinct but overlapping set of ionic currents in each pyloric circuit neuron. Moreover, it can have opposite effects on the same current in different neurons. For example, dopamine has opposite effects on IA in the electrically coupled AB and PD neurons. Adapted from ref. . b, Different neuromodulators influence overlapping but distinct subsets of pyloric circuit neurons. All of the peptidergic and muscarinic actions are excitatory, as are the actions of octopamine. Dopamine and serotonin excite some pyloric neurons and inhibit others. RPCH, red pigment concentrating hormone; LPG, lateral posterior gastric neuron. Peptides and muscarinic actions adapted from ref. ; amines adapted from ref. .
References
-
- Marder E & Calabrese RL Principles of rhythmic motor pattern generation. Physiol. Rev 76, 687–717 (1996). - PubMed
-
- Stein PSG, Grillner S, Selverston AI & Stuart DG (eds) Neurons, Networks, and Motor Behavior (The MIT Press, Cambridge, MA, 1997).
-
- Marder E & Bucher D Central pattern generators and the control of rhythmic movements. Curr. Biol 11, R986–R996 (2001). - PubMed
-
- Pearson KG Neural adaptation in the generation of rhythmic behavior. Annu. Rev. Physiol 62, 723–753 (2000). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
