

Role of phosphatidylinositol(4,5)bisphosphate organization in membrane transport by the Unc104 kinesin motor
- PMID: 12015984
- PMCID: PMC2851634
- DOI: 10.1016/s0092-8674(02)00708-0
Role of phosphatidylinositol(4,5)bisphosphate organization in membrane transport by the Unc104 kinesin motor
Abstract
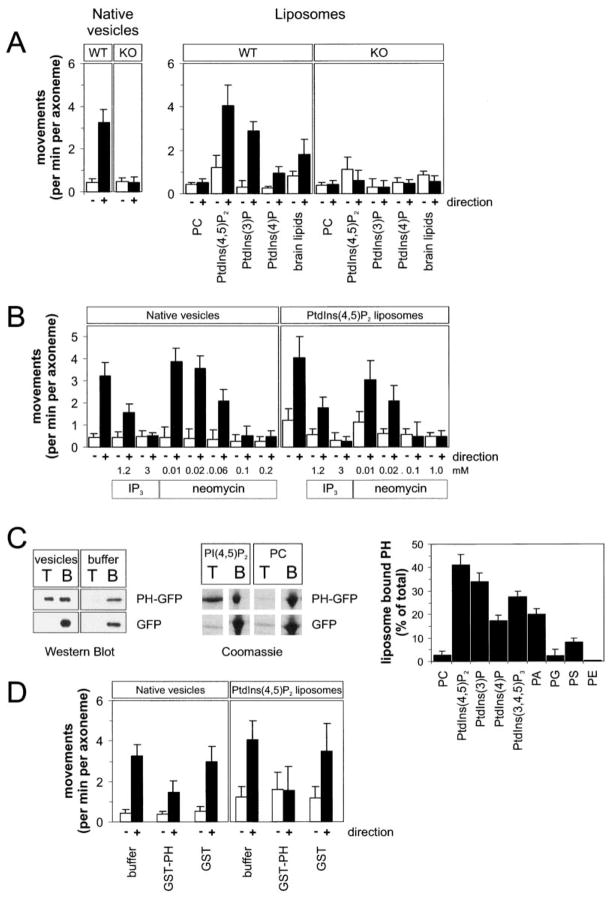
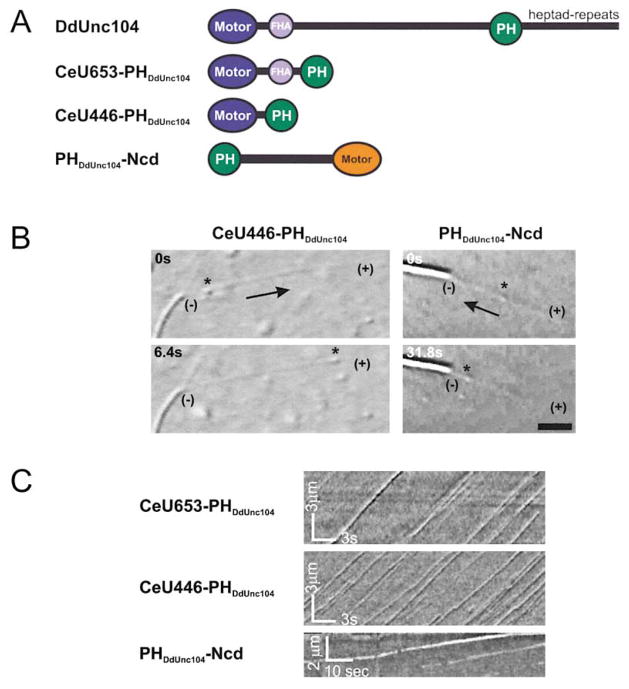
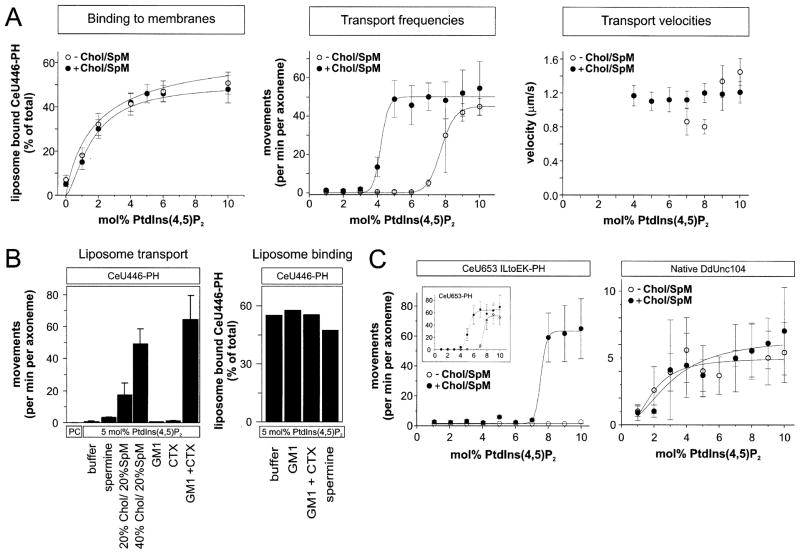
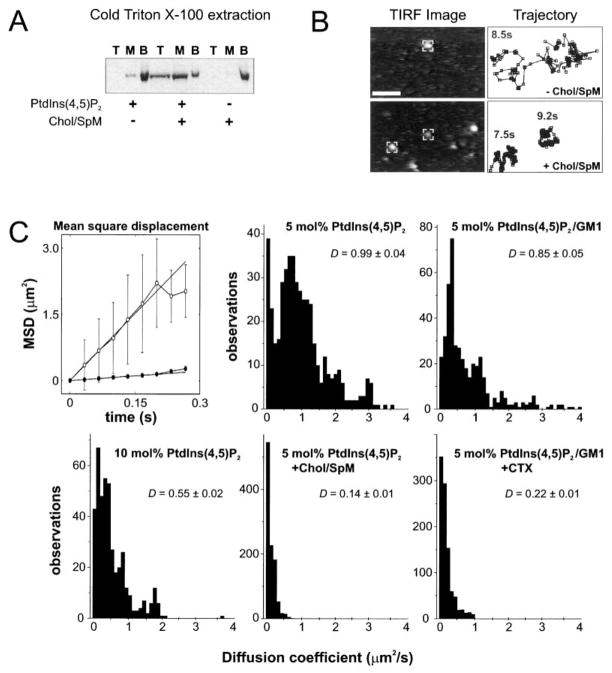
Unc104 (KIF1A) kinesin transports membrane vesicles along microtubules in lower and higher eukaryotes. Using an in vitro motility assay, we show that Unc104 uses a lipid binding pleckstrin homology (PH) domain to dock onto membrane cargo. Through its PH domain, Unc104 can transport phosphatidylinositol(4,5)bisphosphate (PtdIns(4,5)P2)-containing liposomes with similar properties to native vesicles. Interestingly, liposome movement by monomeric Unc104 motors shows a very steep dependence on PtdIns(4,5)P2 concentration (Hill coefficient of approximately 20), even though liposome binding is noncooperative. This switch-like transition for movement can be shifted to lower PtdIns(4,5)P2 concentrations by the addition of cholesterol/sphingomyelin or GM1 ganglioside/cholera toxin, conditions that produce raft-like behavior of Unc104 bound to lipid bilayers. These studies suggest that clustering of Unc104 in PtdIns(4,5)P2-containing rafts provides a trigger for membrane transport.
Figures
References
-
- Berg JS, Derfler BH, Pennisi CM, Corey DP, Cheney RE. Myosin-X, a novel myosin with pleckstrin homology domains, associates with regions of dynamic actin. J Cell Sci. 2000;113:3439–3451. - PubMed
-
- Bloom GS. The UNC-104/KIF1 family of kinesins. Curr Opin Cell Biol. 2001;13:36–40. - PubMed
-
- Case RB, Pierce DW, Hom-Booher N, Hart CL, Vale RD. The directional preference of kinesin motors is specified by an element outside of the motor catalytic domain. Cell. 1997;90:959–966. - PubMed
-
- Cohn RC, Poncz L, Waller RL, Dearborn DG. Phosphoinositide content of erythrocyte membranes in cystic fibrosis. J Lab Clin Med. 1988;111:336–340. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
