

Actin dynamics in platelets
- PMID: 12019562
- PMCID: PMC3376087
- DOI: 10.1016/s0074-7696(02)17014-8
Actin dynamics in platelets
Abstract
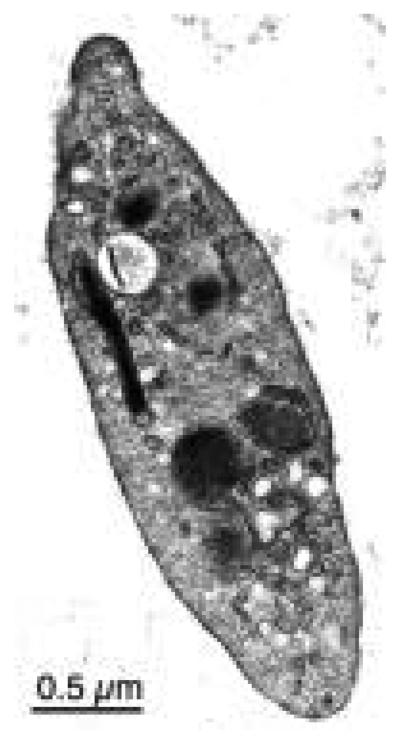
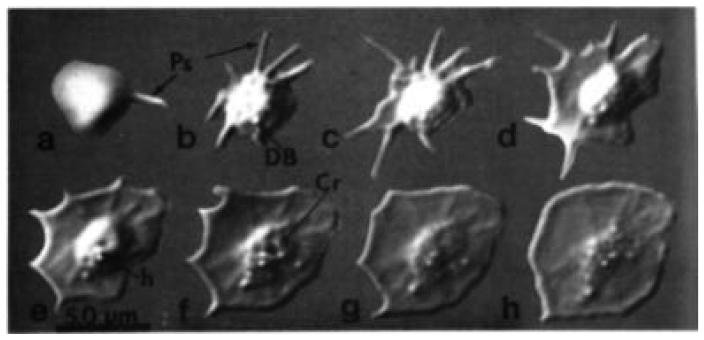
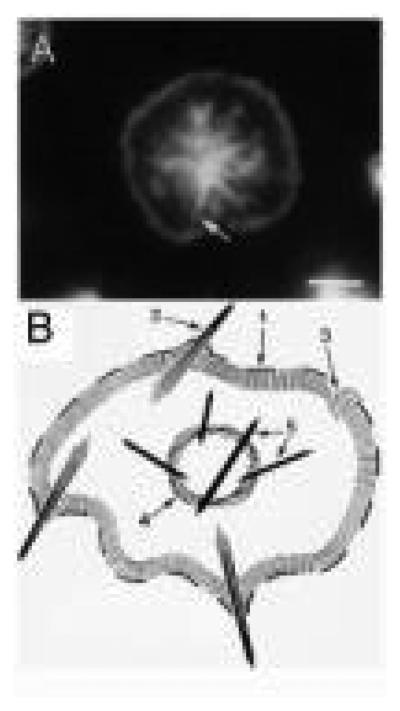
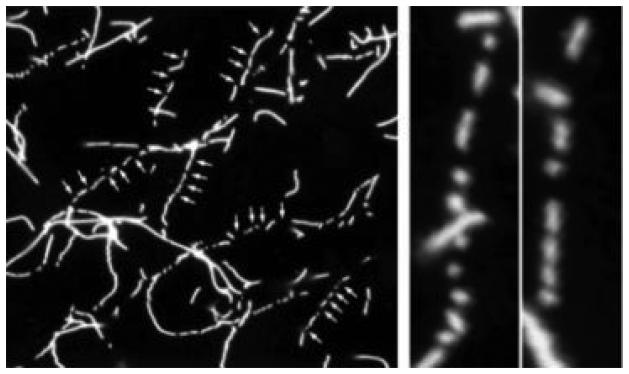
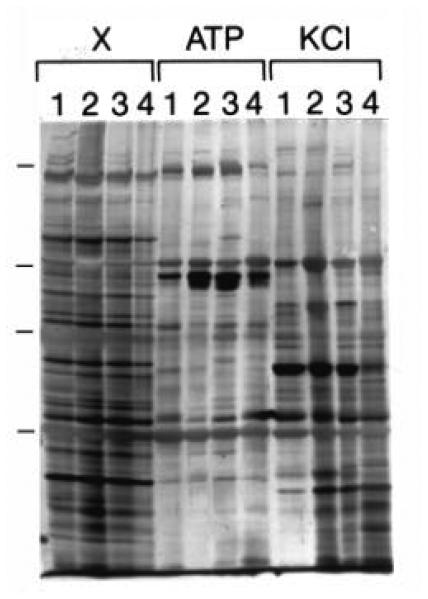
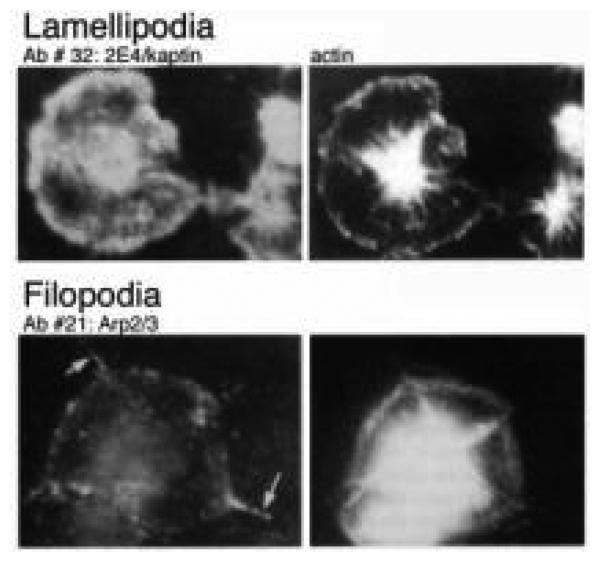
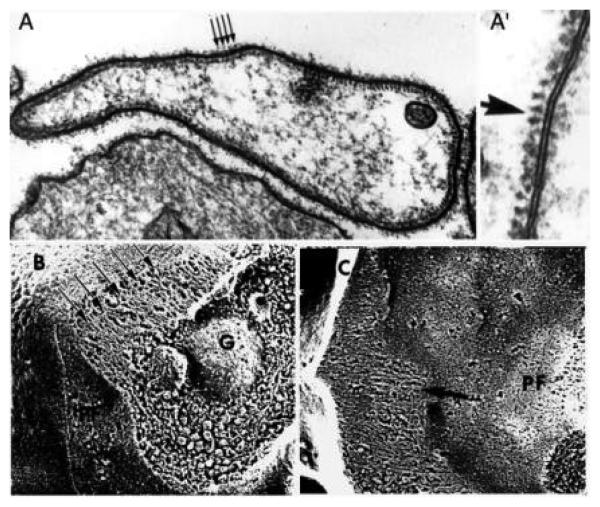
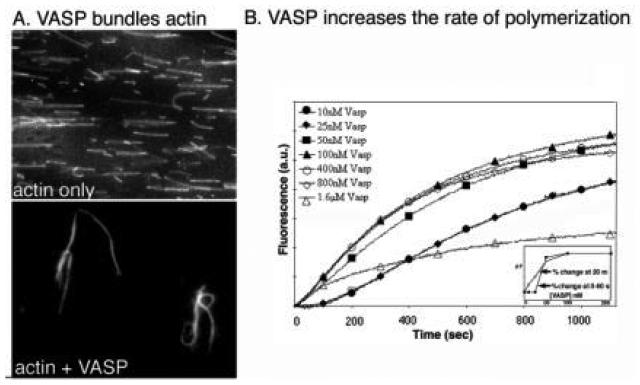
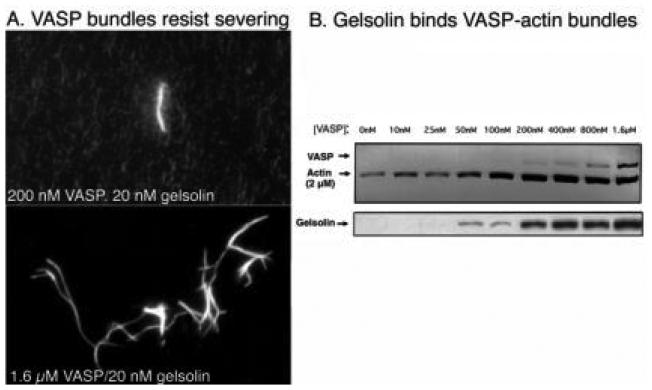
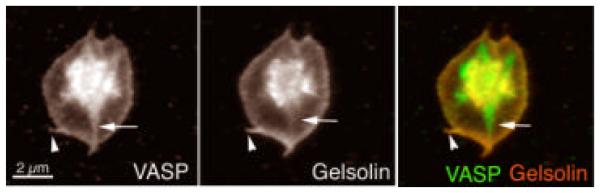
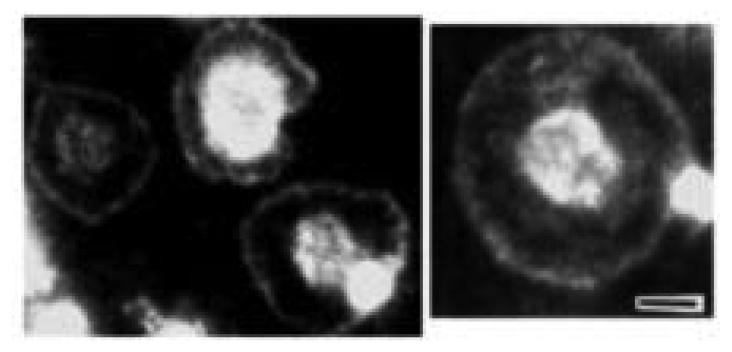
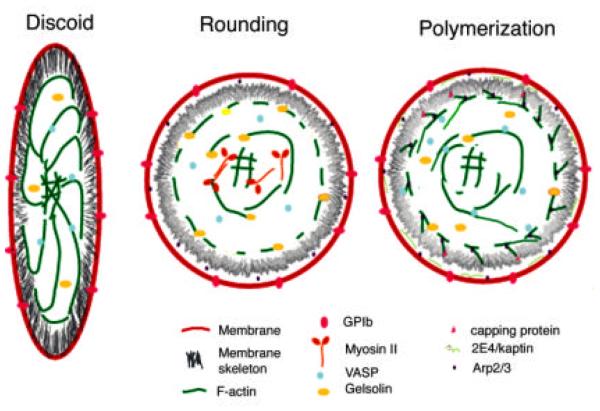
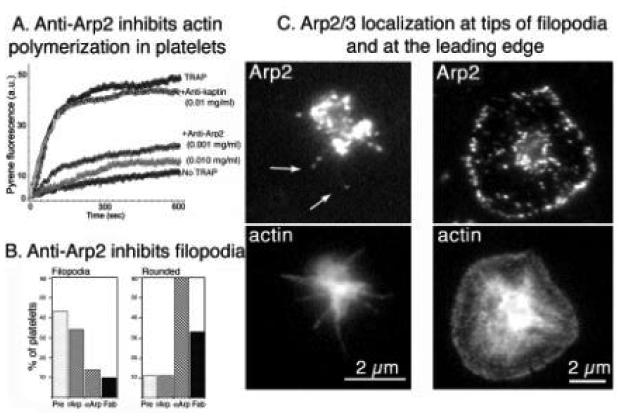
The human blood platelet circulates in the blood as a non-adherent disk. Upon receiving signals of blood vessel damage, the platelet reorganizes its actin cytoskeleton which transforms it into a spiky dynamic adherent glue. This transformation involves a temporal sequence of four morphologically distinct steps which is reproducible in vitro. The actin dynamics underlying these shape changes depend on a large number of actin-binding proteins. Maintenance of the discoid shape requires actin-binding proteins that inhibit these reorganizations, whereas transformation involves other proteins, some to disassemble old filaments and others to polymerize new ones. F-Actin-affinity chromatography identified a large set of actin-binding proteins including VASP, Arp2 and 2E4/kaptin. Recent discoveries show that VASP inhibits filament disassembly and Arp2/3 is required to polymerize new filaments. Morphological analysis of the distribution of these actin-binding proteins in spread platelets together with biochemical measurements of their interactions with actin lead to a model of interactions with actin that mediate shape change.
Figures
References
-
- Abel K, Mieskes G, et al. Dephosphorylation of the focal adhesion protein VASP in vitro and in intact human platelets. FEBS Lett. 1995;370(3):184–8. - PubMed
-
- Amann KJ, Pollard TD. The Arp2/3 complex nucleates actin filament branches from the sides of pre-existing filaments. Nat. Cell Biol. 2001;3(3):306–10. - PubMed
-
- Arber S, Barbayannis FA, et al. Regulation of actin dynamics through phosphorylation of cofilin by LIM-kinase. Nature. 1998;393(6687):805–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources