

Evolution of replication efficiency following infection with a molecularly cloned feline immunodeficiency virus of low virulence
- PMID: 12021339
- PMCID: PMC136200
- DOI: 10.1128/jvi.76.12.6062-6072.2002
Evolution of replication efficiency following infection with a molecularly cloned feline immunodeficiency virus of low virulence
Abstract
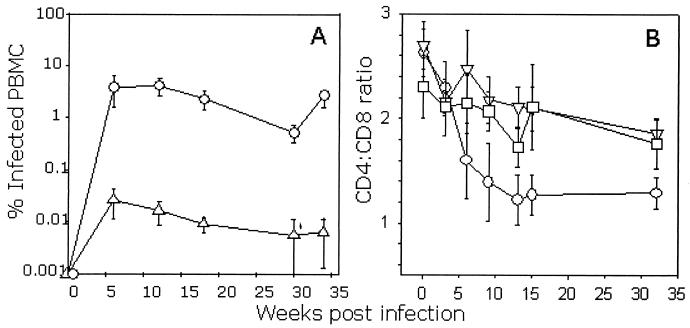
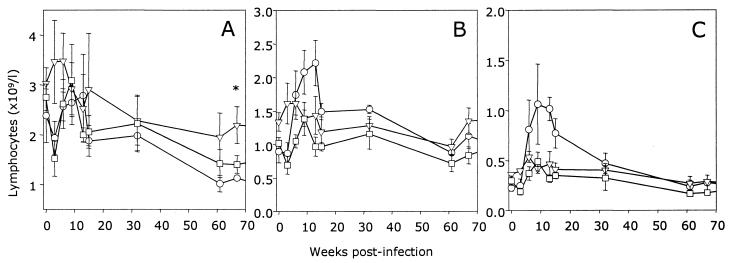
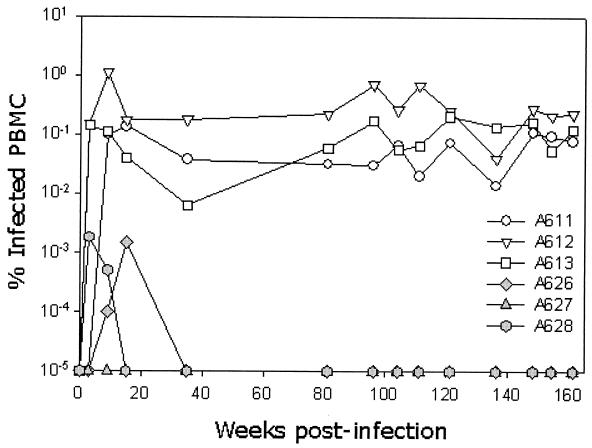
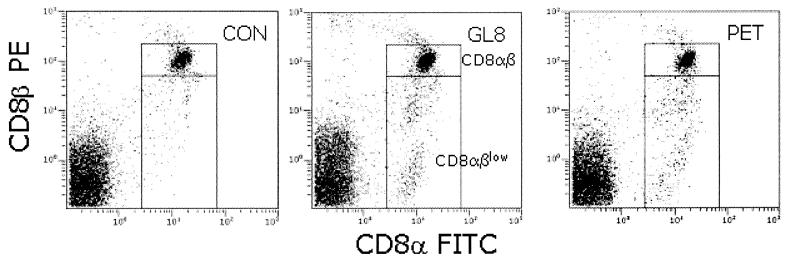
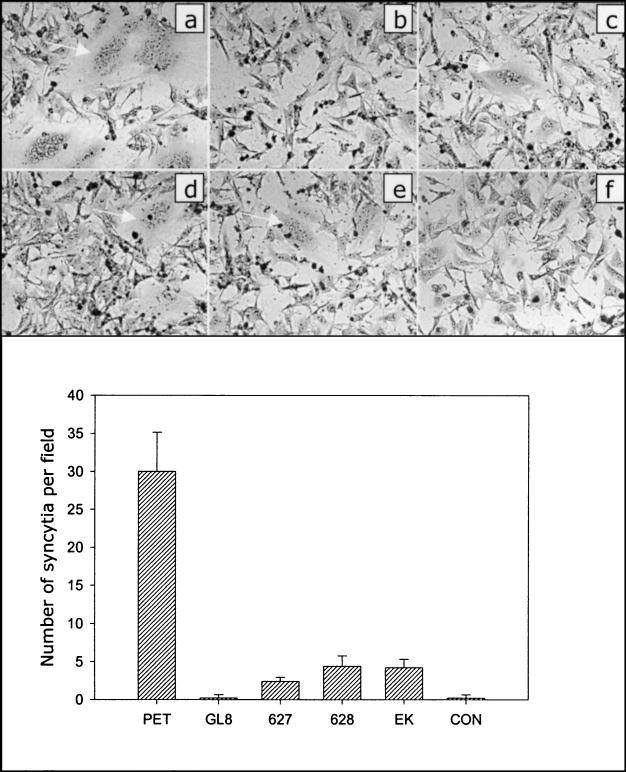
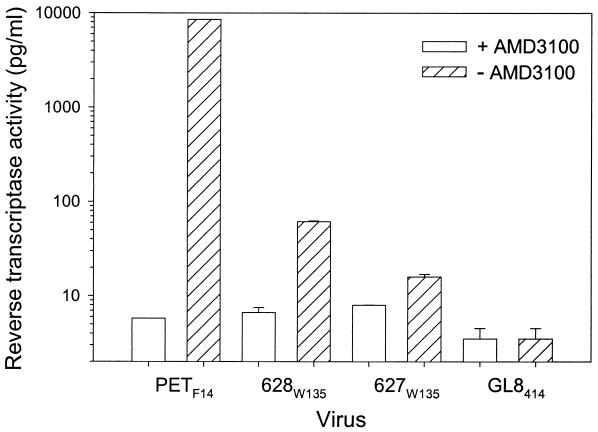
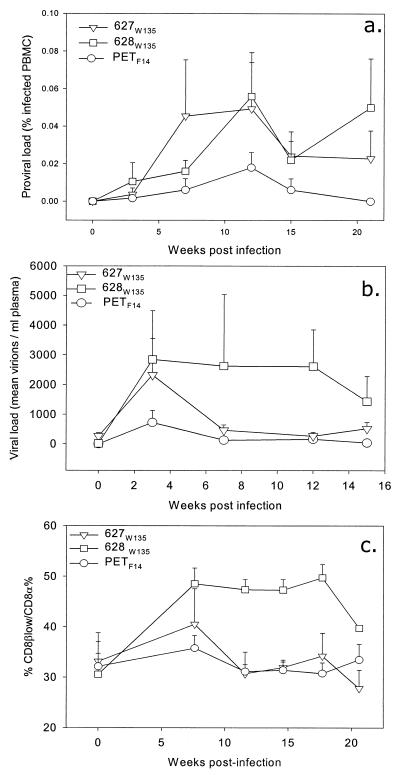
The development of an effective vaccine against human immunodeficiency virus is considered to be the most practicable means of controlling the advancing global AIDS epidemic. Studies with the domestic cat have demonstrated that vaccinal immunity to infection can be induced against feline immunodeficiency virus (FIV); however, protection is largely restricted to laboratory strains of FIV and does not extend to primary strains of the virus. We compared the pathogenicity of two prototypic vaccine challenge strains of FIV derived from molecular clones; the laboratory strain PET(F14) and the primary strain GL8(414). PET(F14) established a low viral load and had no effect on CD4(+)- or CD8(+)-lymphocyte subsets. In contrast, GL8(414) established a high viral load and induced a significant reduction in the ratio of CD4(+) to CD8(+) lymphocytes by 15 weeks postinfection, suggesting that PET(F14) may be a low-virulence-challenge virus. However, during long-term monitoring of the PET(F14)-infected cats, we observed the emergence of variant viruses in two of three cats. Concomitant with the appearance of the variant viruses, designated 627(W135) and 628(W135,) we observed an expansion of CD8(+)-lymphocyte subpopulations expressing reduced CD8 beta-chain, a phenotype consistent with activation. The variant viruses both carried mutations that reduced the net charge of the V3 loop (K409Q and K409E), giving rise to a reduced ability of the Env proteins to both induce fusion and to establish productive infection in CXCR4-expressing cells. Further, following subsequent challenge of naïve cats with the mutant viruses, the viruses established higher viral loads and induced more marked alterations in CD8(+)-lymphocyte subpopulations than did the parent F14 strain of virus, suggesting that the E409K mutation in the PET(F14) strain contributes to the attenuation of the virus.
Figures

Similar articles
-
Feline immunodeficiency virus (FIV) neutralization: a review.Viruses. 2011 Oct;3(10):1870-90. doi: 10.3390/v3101870. Epub 2011 Oct 13. Viruses. 2011. PMID: 22069520 Free PMC article. Review.
-
Feline immunodeficiency virus xenoinfection: the role of chemokine receptors and envelope diversity.J Virol. 2002 Apr;76(8):3626-36. doi: 10.1128/jvi.76.8.3626-3636.2002. J Virol. 2002. PMID: 11907202 Free PMC article.
-
Feline immunodeficiency virus env gene evolution in experimentally infected cats.Vet Immunol Immunopathol. 2010 Mar 15;134(1-2):96-106. doi: 10.1016/j.vetimm.2009.10.015. Epub 2009 Nov 7. Vet Immunol Immunopathol. 2010. PMID: 19897254
-
Prior virus exposure alters the long-term landscape of viral replication during feline lentiviral infection.Viruses. 2011 Oct;3(10):1891-908. doi: 10.3390/v3101891. Epub 2011 Oct 13. Viruses. 2011. PMID: 22069521 Free PMC article.
-
Early events in the immunopathogenesis of feline retrovirus infections.J Am Vet Med Assoc. 1991 Nov 15;199(10):1311-5. J Am Vet Med Assoc. 1991. PMID: 1666073 Review.
Cited by
-
Characterization of a highly pathogenic molecular clone of feline immunodeficiency virus clade C.J Virol. 2004 Sep;78(17):8971-82. doi: 10.1128/JVI.78.17.8971-8982.2004. J Virol. 2004. PMID: 15308694 Free PMC article.
-
A single site for N-linked glycosylation in the envelope glycoprotein of feline immunodeficiency virus modulates the virus-receptor interaction.Retrovirology. 2008 Aug 22;5:77. doi: 10.1186/1742-4690-5-77. Retrovirology. 2008. PMID: 18721458 Free PMC article.
-
The molecular biology of feline immunodeficiency virus (FIV).Viruses. 2011 Nov;3(11):2192-213. doi: 10.3390/v3112192. Epub 2011 Nov 9. Viruses. 2011. PMID: 22163340 Free PMC article. Review.
-
Dynamics of viral and proviral loads of feline immunodeficiency virus within the feline central nervous system during the acute phase following intravenous infection.J Virol. 2003 Jul;77(13):7477-85. doi: 10.1128/jvi.77.13.7477-7485.2003. J Virol. 2003. PMID: 12805447 Free PMC article.
-
Feline immunodeficiency virus (FIV) neutralization: a review.Viruses. 2011 Oct;3(10):1870-90. doi: 10.3390/v3101870. Epub 2011 Oct 13. Viruses. 2011. PMID: 22069520 Free PMC article. Review.
References
-
- Bachmann, M. H., C. Mathiason-Dubard, G. H. Learn, A. G. Rodrigo, D. L. Sodora, P. Mazzetti, E. A. Hoover, and J. I. Mullins. 1997. Genetic diversity of feline immunodeficiency virus: dual infection, recombination, and distinct evolutionary rates among envelope sequence clades. J. Virol. 71:4241-4253. - PMC - PubMed
-
- Bendinelli, M., M. Pistello, D. Del Mauro, G. Cammarota, F. Maggi, A. Leonildi, S. Giannecchini, C. Bergamini, and D. Matteucci. 2001. During readaptation in vivo, a tissue culture-adapted strain of feline immunodeficiency virus reverts to broad neutralization resistance at different times in individual hosts but through changes at the same position of the surface glycoprotein. J. Virol. 75:4584-4593. - PMC - PubMed
-
- Cammarota, G., D. Matteucci, M. Pistello, E. Nicoletti, S. Giannecchini, and M. Bendinelli. 1996. Reduced sensitivity to strain-specific neutralization of laboratory-adapted feline immunodeficiency virus after one passage in vivo: association with amino acid substitutions in the V4 region of the surface glycoprotein. AIDS Res. Hum. Retrovir. 12:173-175. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
