

Disulfide bond configuration of human cytomegalovirus glycoprotein B
- PMID: 12021340
- PMCID: PMC136243
- DOI: 10.1128/jvi.76.12.6073-6082.2002
Disulfide bond configuration of human cytomegalovirus glycoprotein B
Abstract
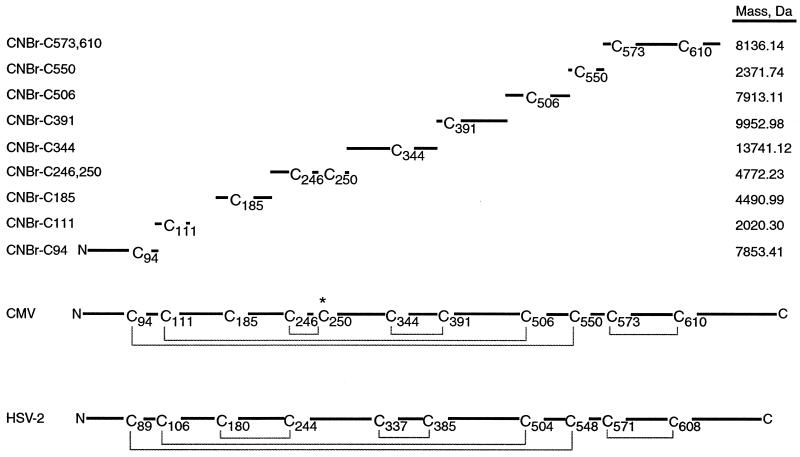
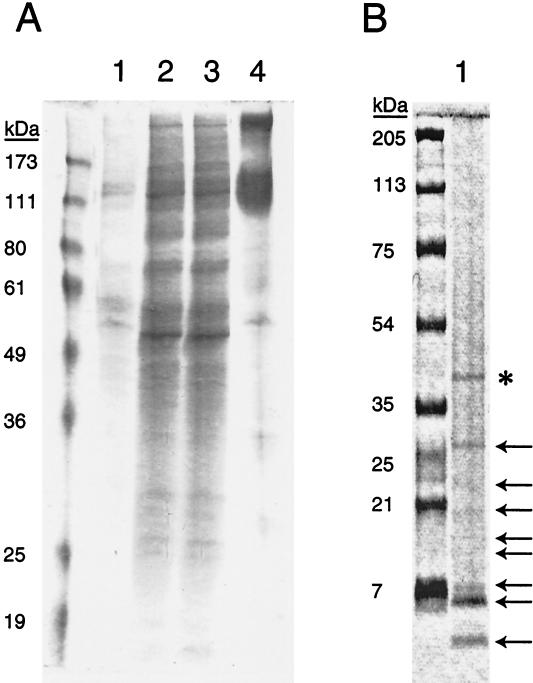
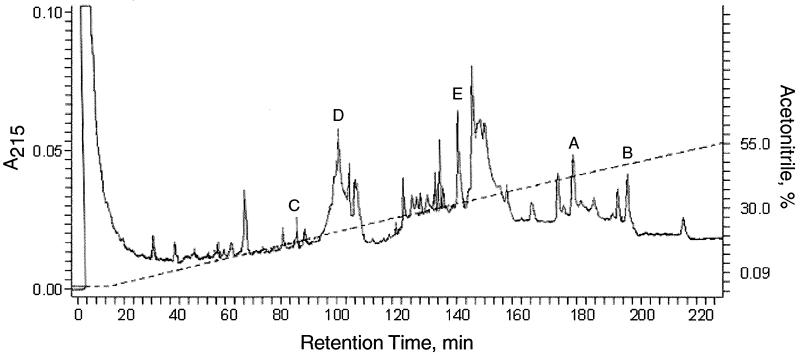
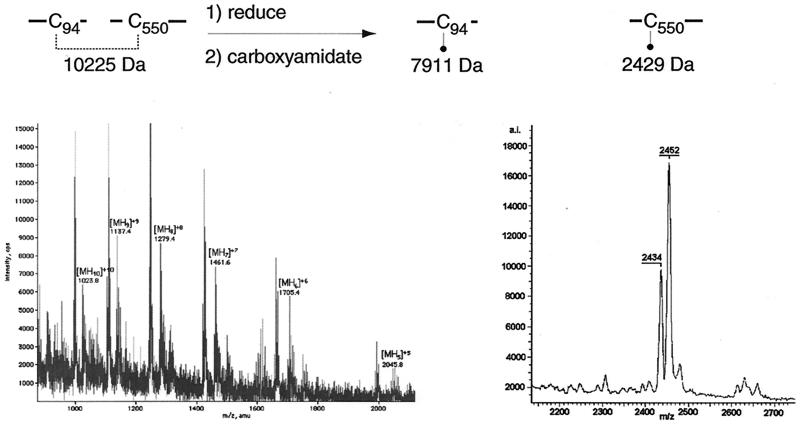
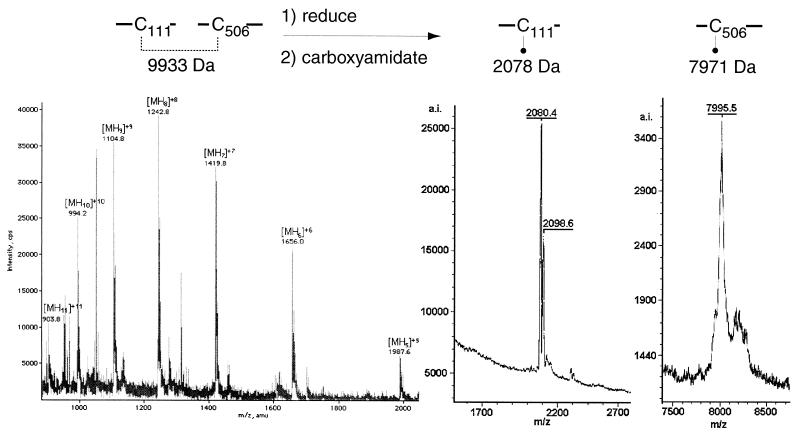
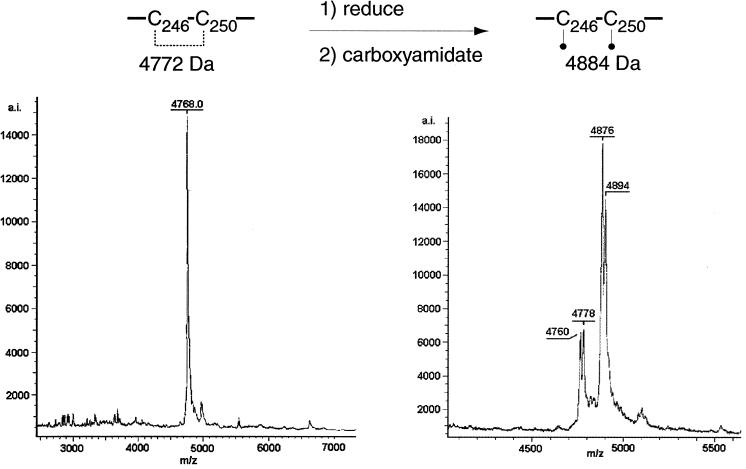
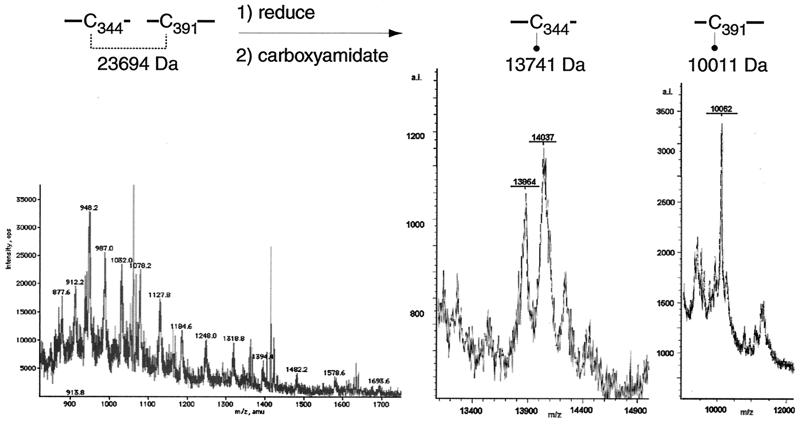
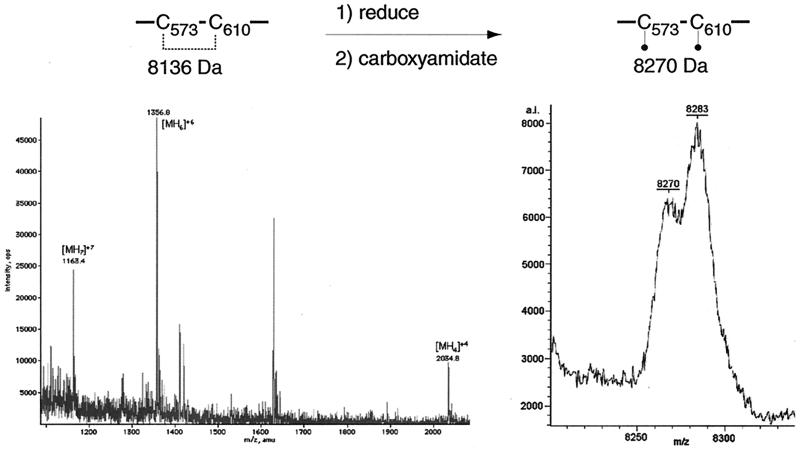
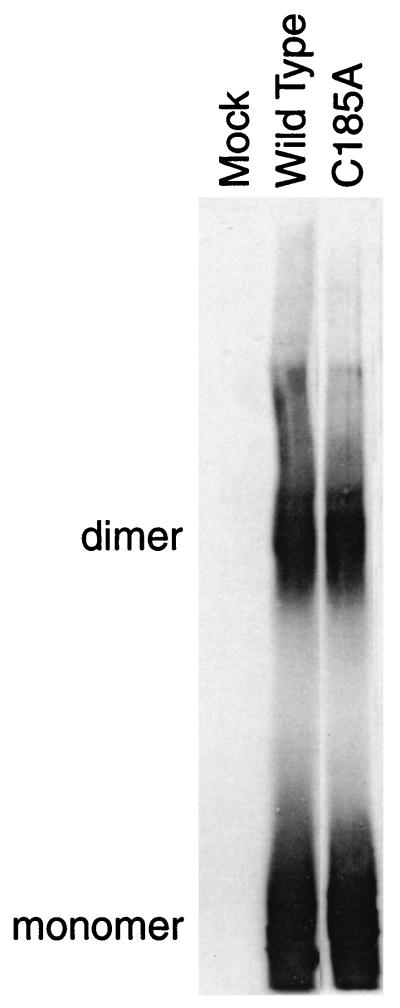
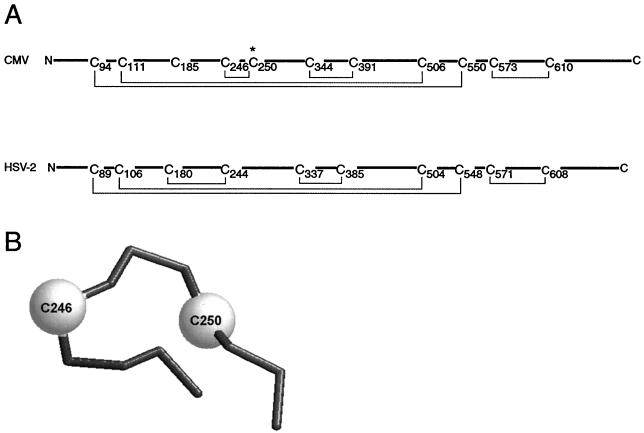
Glycoprotein B (gB) is the most highly conserved of the envelope glycoproteins of human herpesviruses. The gB protein of human cytomegalovirus (CMV) serves multiple roles in the life cycle of the virus. To investigate structural properties of gB that give rise to its function, we sought to determine the disulfide bond arrangement of gB. To this end, a recombinant form of gB (gB-S) comprising the entire ectodomain of the glycoprotein (amino acids 1 to 750) was constructed and expressed in insect cells. Proteolytic fragmentation and mass spectrometry were performed using purified gB-S, and the five disulfide bonds that link 10 of the 11 highly conserved cysteine residues of gB were mapped. These bonds are C94-C550, C111-C506, C246-C250, C344-C391, and C573-C610. This configuration closely parallels the disulfide bond configuration of herpes simplex type 2 (HSV-2) gB (N. Norais, D. Tang, S. Kaur, S. H. Chamberlain, F. R. Masiarz, R. L. Burke, and F. Markus, J. Virol. 70:7379-7387, 1996). However, despite the high degree of conservation of cysteine residues between CMV gB and HSV-2 gB, the disulfide bond arrangements of the two homologs are not identical. We detected a disulfide bond between the conserved cysteine residue 246 and the nonconserved cysteine residue 250 of CMV gB. We hypothesize that this disulfide bond stabilizes a tight loop in the amino-terminal fragment of CMV gB that does not exist in HSV-2 gB. We predicted that the cysteine residue not found in a disulfide bond of CMV gB, cysteine residue 185, would play a role in dimerization, but a cysteine substitution mutant in cysteine residue 185 showed no apparent defect in the ability to form dimers. These results indicate that gB oligomerization involves additional interactions other than a single disulfide bond. This work represents the second reported disulfide bond structure for a herpesvirus gB homolog, and the discovery that the two structures are not identical underscores the importance of empirically determining structures even for highly conserved proteins.
Figures
References
-
- Britt, W. J. 1984. Neutralizing antibodies detect a disulfide-linked glycoprotein complex within the envelope of human cytomegalovirus. Virology 135:369-378. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
