

Subcellular localization and integration activities of rous sarcoma virus reverse transcriptase
- PMID: 12021354
- PMCID: PMC136205
- DOI: 10.1128/jvi.76.12.6205-6212.2002
Subcellular localization and integration activities of rous sarcoma virus reverse transcriptase
Abstract
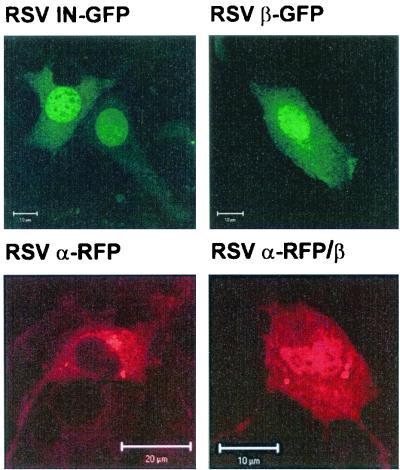
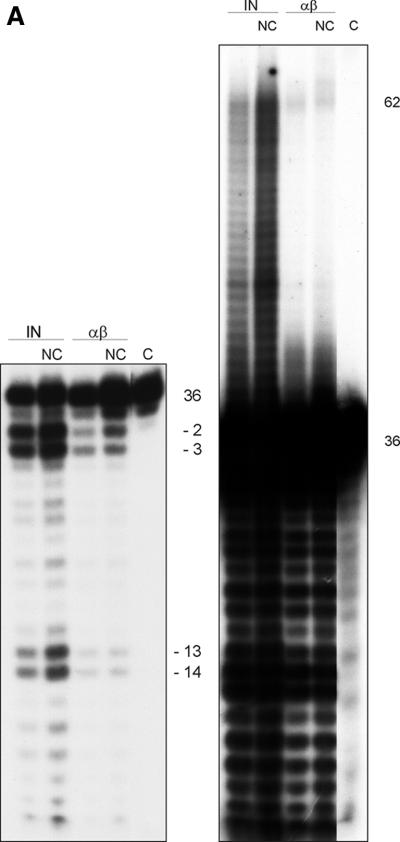
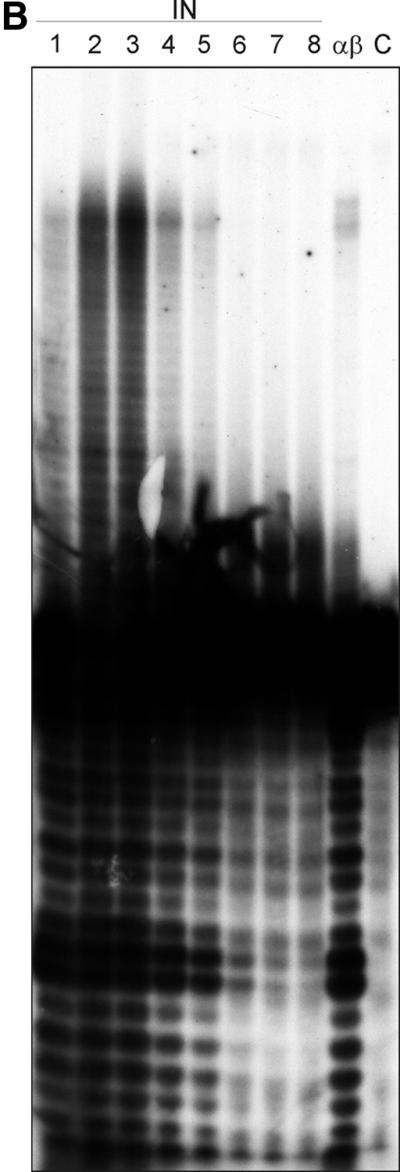
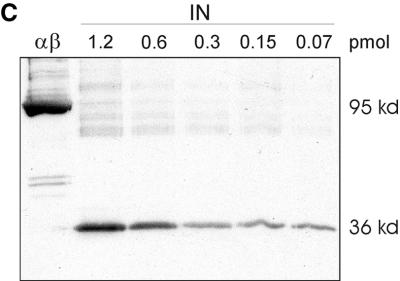
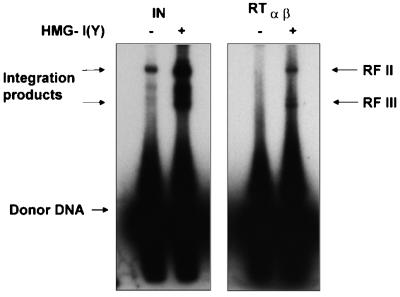
Reverse transcriptases (RTs) alphabeta and beta from avian Rous sarcoma virus (RSV) harbor an integrase domain which is absent in nonavian retroviral RTs. RSV integrase contains a nuclear localization signal which enables the enzyme to enter the nucleus of the cell in order to perform integration of the proviral DNA into the host genome. In the present study we analyzed the subcellular localization of RSV RT, since previous results indicated that RSV finishes synthesis of the proviral DNA in the nucleus. Our results demonstrate that the heterodimeric RSV RT alphabeta and the beta subunit, when expressed independently, can be detected in the nucleus, whereas the separate alpha subunit lacking the integrase domain is prevalent in the cytoplasm. These data suggest an involvement of RSV RT in the transport of the preintegration complex into the nucleus. In addition, to analyze whether the integrase domain, located at the carboxyl terminus of beta, exhibits integration activities, we investigated the nicking and joining activities of heterodimeric RSV RT alphabeta with an oligodeoxynucleotide-based assay system and with a donor substrate containing the supF gene flanked by the viral long terminal repeats. Our data show that RSV RT alphabeta is able to perform the integration reaction in vitro; however, it does so with an estimated 30-fold lower efficiency than the free RSV integrase, indicating that RSV RT is not involved in integration in vivo. Integration with RSV RT alphabeta could be stimulated in the presence of human immunodeficiency virus type 1 nucleocapsid protein or HMG-I(Y).
Figures

Similar articles
-
Soluble Rous sarcoma virus reverse transcriptases alpha, alphabeta, and beta purified from insect cells are processive DNA polymerases that lack an RNase H 3' --> 5' directed processing activity.J Biol Chem. 1999 Sep 10;274(37):26329-36. doi: 10.1074/jbc.274.37.26329. J Biol Chem. 1999. PMID: 10473589
-
Molecular and genetic determinants of rous sarcoma virus integrase for concerted DNA integration.J Virol. 2003 Jun;77(11):6482-92. doi: 10.1128/jvi.77.11.6482-6492.2003. J Virol. 2003. PMID: 12743305 Free PMC article.
-
Rous sarcoma virus integrase protein: mapping functions for catalysis and substrate binding.J Virol. 1994 Apr;68(4):2215-23. doi: 10.1128/JVI.68.4.2215-2223.1994. J Virol. 1994. PMID: 8139006 Free PMC article.
-
Oligomerization of Retrovirus Integrases.Subcell Biochem. 2018;88:211-243. doi: 10.1007/978-981-10-8456-0_10. Subcell Biochem. 2018. PMID: 29900499 Review.
-
Retroviral DNA integration.Microbiol Mol Biol Rev. 1999 Dec;63(4):836-43, table of contents. doi: 10.1128/MMBR.63.4.836-843.1999. Microbiol Mol Biol Rev. 1999. PMID: 10585967 Free PMC article. Review.
Cited by
-
Cooperation between reverse transcriptase and integrase during reverse transcription and formation of the preintegrative complex of Ty1.Eukaryot Cell. 2006 Oct;5(10):1760-9. doi: 10.1128/EC.00159-06. Eukaryot Cell. 2006. PMID: 17031000 Free PMC article.
-
Equine infectious anemia virus Gag p9 function in early steps of virus infection and provirus production.J Virol. 2005 Jul;79(14):8793-801. doi: 10.1128/JVI.79.14.8793-8801.2005. J Virol. 2005. PMID: 15994773 Free PMC article.
-
Nuclear trafficking of retroviral RNAs and Gag proteins during late steps of replication.Viruses. 2013 Nov 18;5(11):2767-95. doi: 10.3390/v5112767. Viruses. 2013. PMID: 24253283 Free PMC article. Review.
-
Nuclear import of Avian Sarcoma Virus integrase is facilitated by host cell factors.Retrovirology. 2008 Aug 7;5:73. doi: 10.1186/1742-4690-5-73. Retrovirology. 2008. PMID: 18687138 Free PMC article.
-
Role of integrase in reverse transcription of the Saccharomyces cerevisiae retrotransposon Ty1.Eukaryot Cell. 2005 Jun;4(6):1057-65. doi: 10.1128/EC.4.6.1057-1065.2005. Eukaryot Cell. 2005. PMID: 15947198 Free PMC article.
References
-
- Baltimore, D. 1970. RNA-dependent DNA polymerase in virions of RNA tumor viruses. Nature 226:1209-1211. - PubMed
-
- Bowerman, B., P. O. Brown, J. M. Bishop, and H. E. Varmus. 1989. A nucleoprotein complex mediates the integration of retroviral DNA. Genes Dev. 3:469-478. - PubMed
-
- Brin, E., J. Yi, A. M. Skalka, and J. Leis. 2000. Modeling the late steps in HIV-1 retroviral integrase-catalyzed DNA integration. J. Biol. Chem. 275:39287-39295. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
