

Structural basis of alpha-amylase activation by chloride
- PMID: 12021442
- PMCID: PMC2373612
- DOI: 10.1110/ps.0202602
Structural basis of alpha-amylase activation by chloride
Abstract
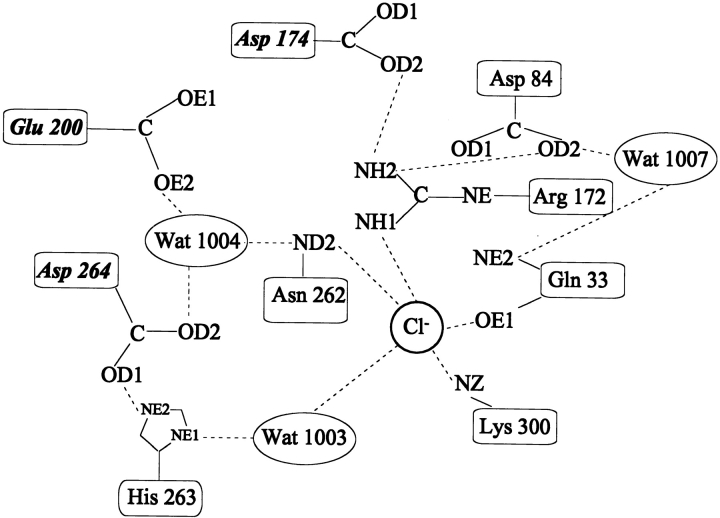
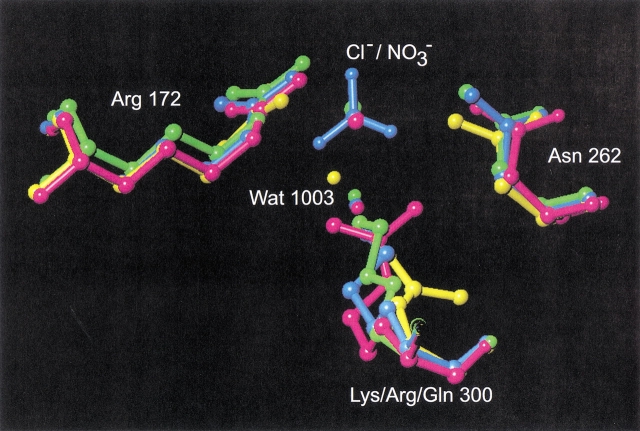
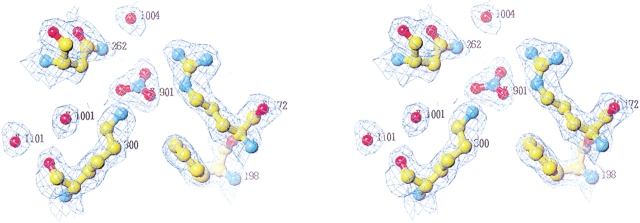
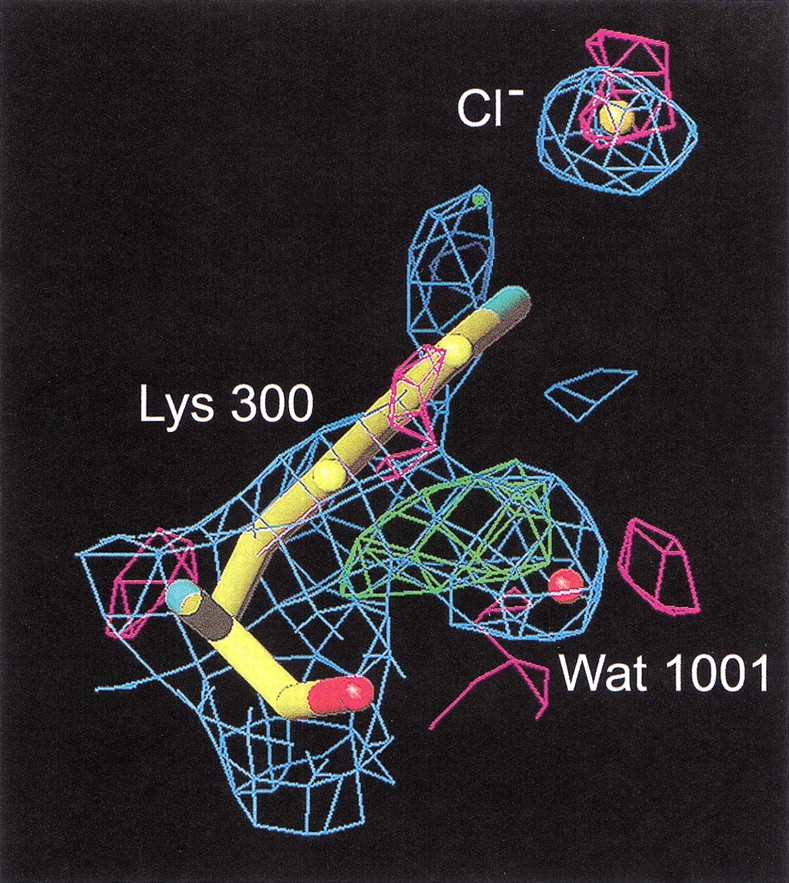
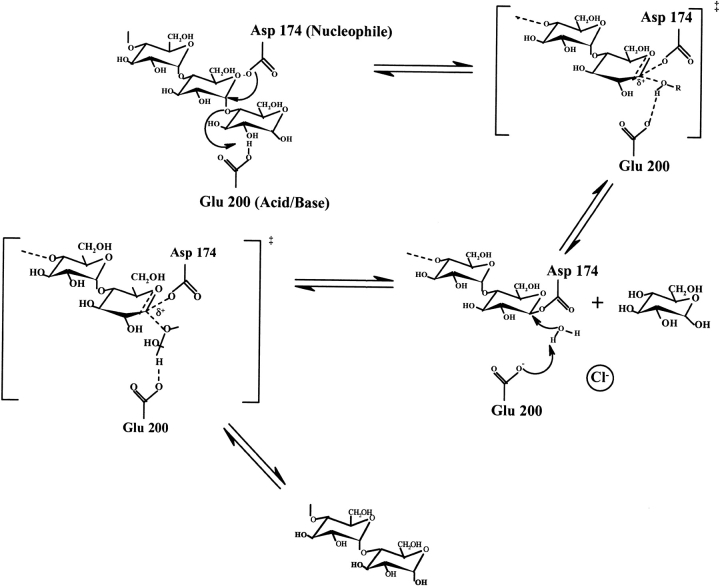
To further investigate the mechanism and function of allosteric activation by chloride in some alpha-amylases, the structure of the bacterial alpha-amylase from the psychrophilic micro-organism Pseudoalteromonas haloplanktis in complex with nitrate has been solved at 2.1 A degrees, as well as the structure of the mutants Lys300Gln (2.5 A degrees ) and Lys300Arg (2.25 A degrees ). Nitrate binds strongly to alpha-amylase but is a weak activator. Mutation of the critical chloride ligand Lys300 into Gln results in a chloride-independent enzyme, whereas the mutation into Arg mimics the binding site as is found in animal alpha-amylases with, however, a lower affinity for chloride. These structures reveal that the triangular conformation of the chloride ligands and the nearly equatorial coordination allow the perfect accommodation of planar trigonal monovalent anions such as NO3-, explaining their unusual strong binding. It is also shown that a localized negative charge such as that of Cl-, rather than a delocalized charge as in the case of nitrate, is essential for maximal activation. The chloride-free mutant Lys300Gln indicates that chloride is not mandatory for the catalytic mechanism but strongly increases the reactivity at the active site. Disappearance of the putative catalytic water molecule in this weakly active mutant supports the view that chloride helps to polarize the hydrolytic water molecule and enhances the rate of the second step in the catalytic reaction.
Figures
References
-
- ———. 1998b. Structures of the psychrophilic Alteromonas haloplanctis α-amylase give insights into cold adaptation at a molecular level. Structure 6 1503–1516. - PubMed
-
- Aghajari, N., Roth, M., and Haser, R. 2002. Crystallographic evidence of a transglycosylation reaction: Ternary complexes of a psychrophilic α-amylase. Biochemistry 41 4273–4280. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
