

Photoexcited structure of a plant photoreceptor domain reveals a light-driven molecular switch
- PMID: 12034897
- PMCID: PMC150607
- DOI: 10.1105/tpc.010475
Photoexcited structure of a plant photoreceptor domain reveals a light-driven molecular switch
Abstract
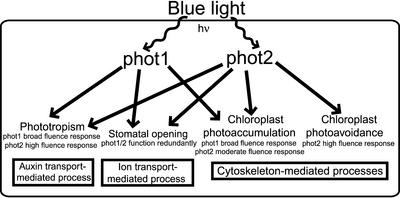
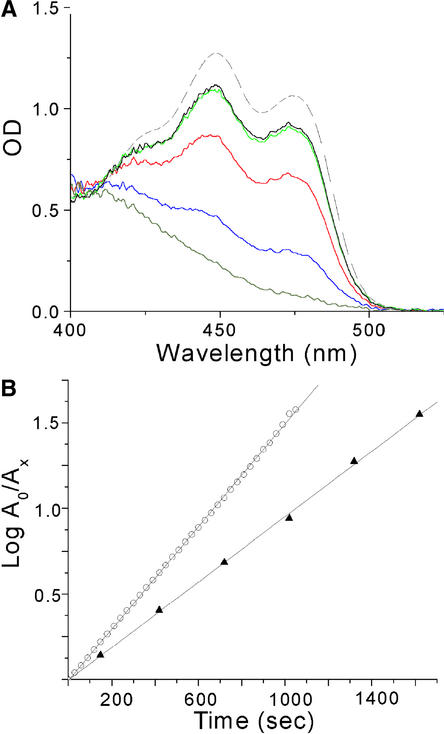
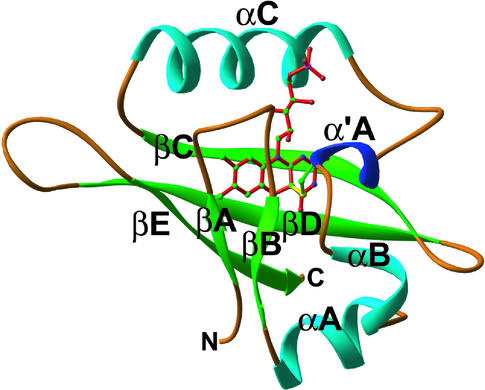
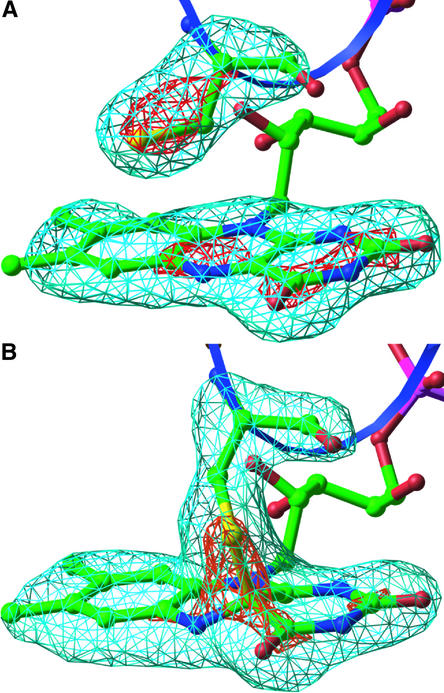
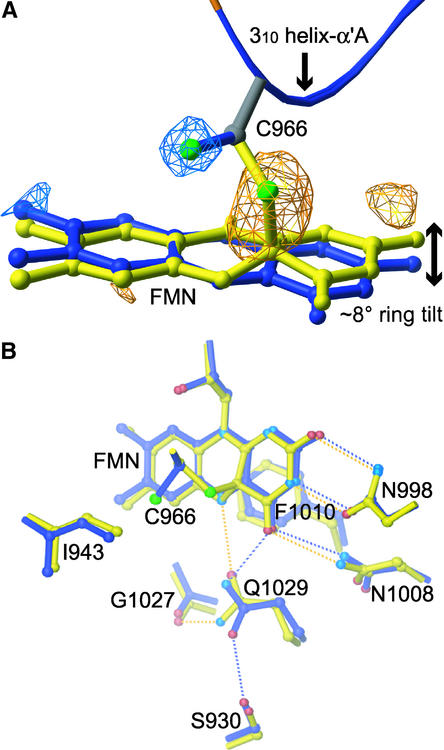
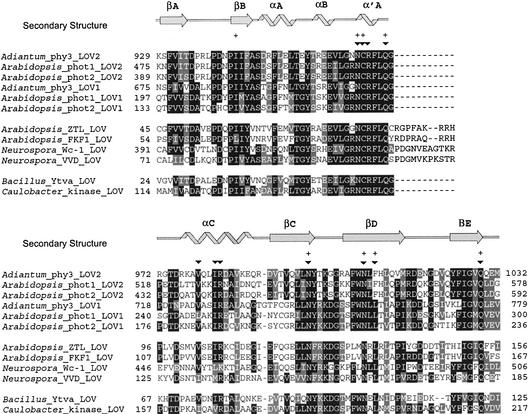
The phototropins are flavoprotein kinases that control phototropic bending, light-induced chloroplast movement, and stomatal opening in plants. Two flavin mononucleotide binding light, oxygen, or voltage (LOV) domains are the sites for initial photochemistry in these blue light photoreceptors. We have determined the steady state, photoexcited crystal structure of a flavin-bound LOV domain. The structure reveals a unique photochemical switch in the flavin binding pocket in which the absorption of light drives the formation of a reversible covalent bond between a highly conserved Cys residue and the flavin cofactor. This provides a molecular picture of a cysteinyl-flavin covalent adduct, the presumed signaling species that leads to phototropin kinase activation and subsequent signal transduction. We identify closely related LOV domains in two eubacterial proteins that suggests the light-induced conformational change evident in this structure is an ancient biomolecular response to light, arising before the appearance of plants.
Figures
References
-
- Arabidopsis Genome Initiative. (2000). Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408, 796–815. - PubMed
-
- Bailey, S. (1994). The CCP4 suite: Programs for protein crystallography. Acta Crystallogr. D 50, 760–763. - PubMed
-
- Bolognesi, M., Ghisla, S., and Incoccia, L. (1978). Crystal and molecular structure of 2 models of catalytic flavo(co)enzyme intermediates. Acta Crystallogr. B 34, 821–828.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
