

Arp2/3 complex is required for actin polymerization during platelet shape change
- PMID: 12036877
- PMCID: PMC3376088
- DOI: 10.1182/blood.v99.12.4466
Arp2/3 complex is required for actin polymerization during platelet shape change
Abstract
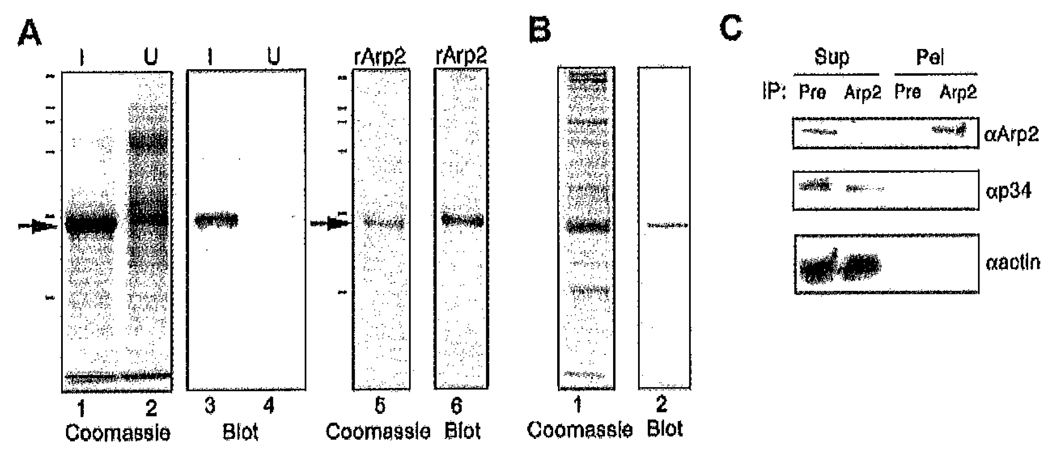
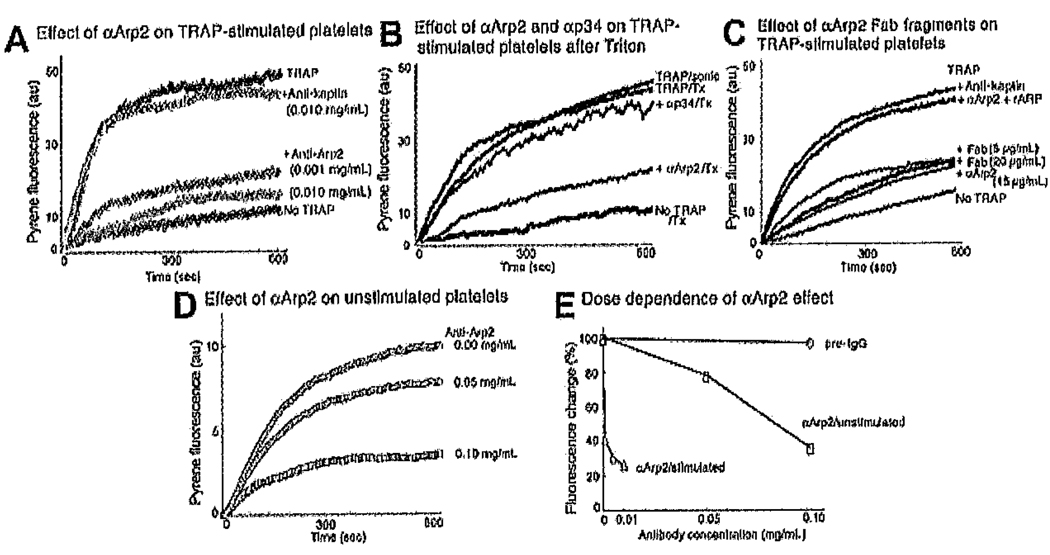
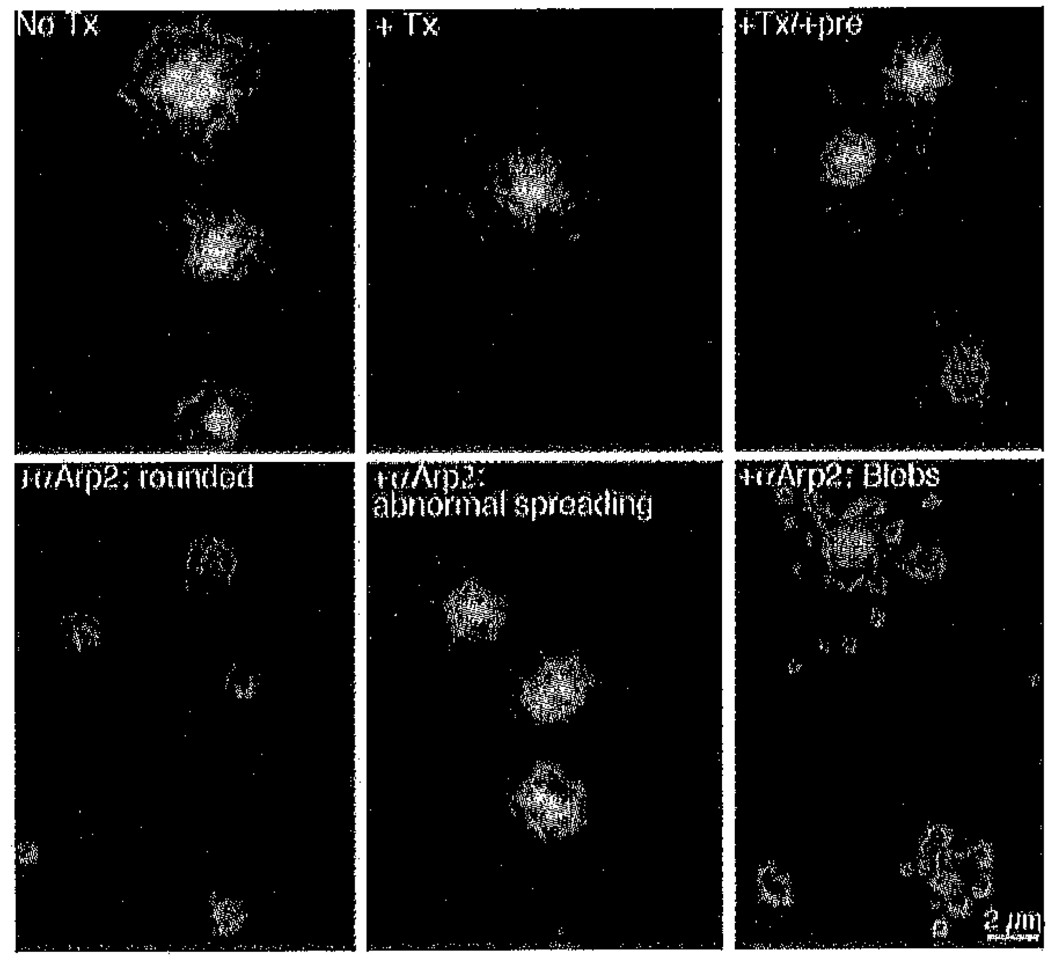
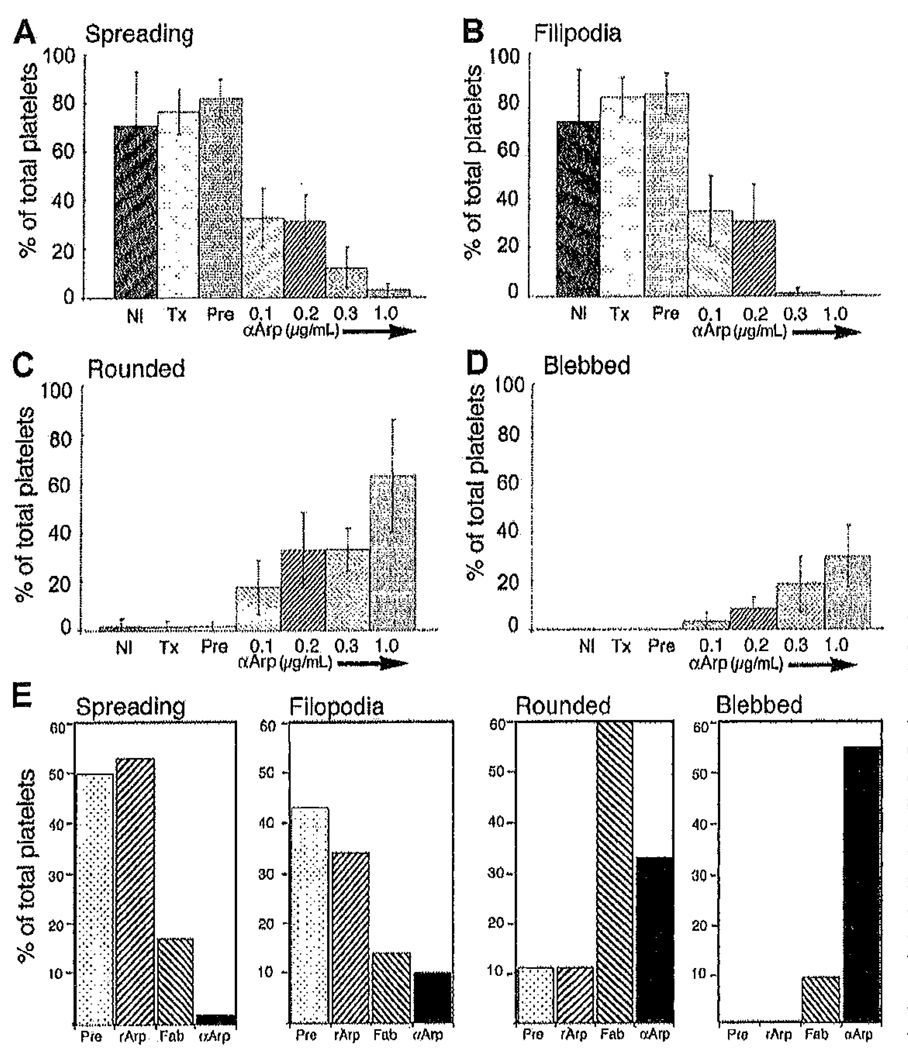
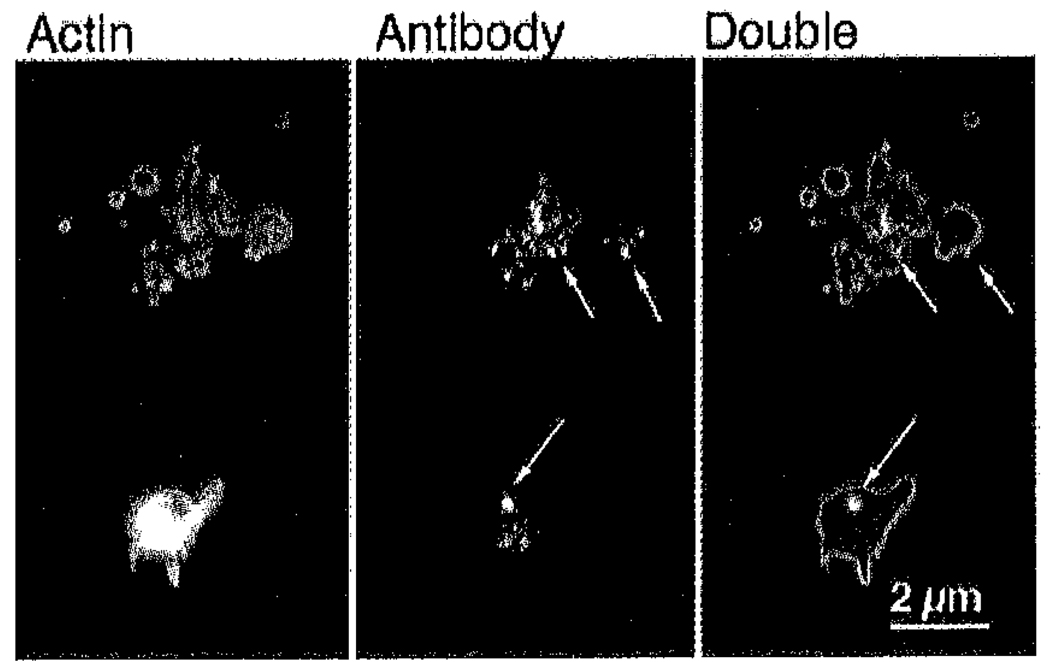
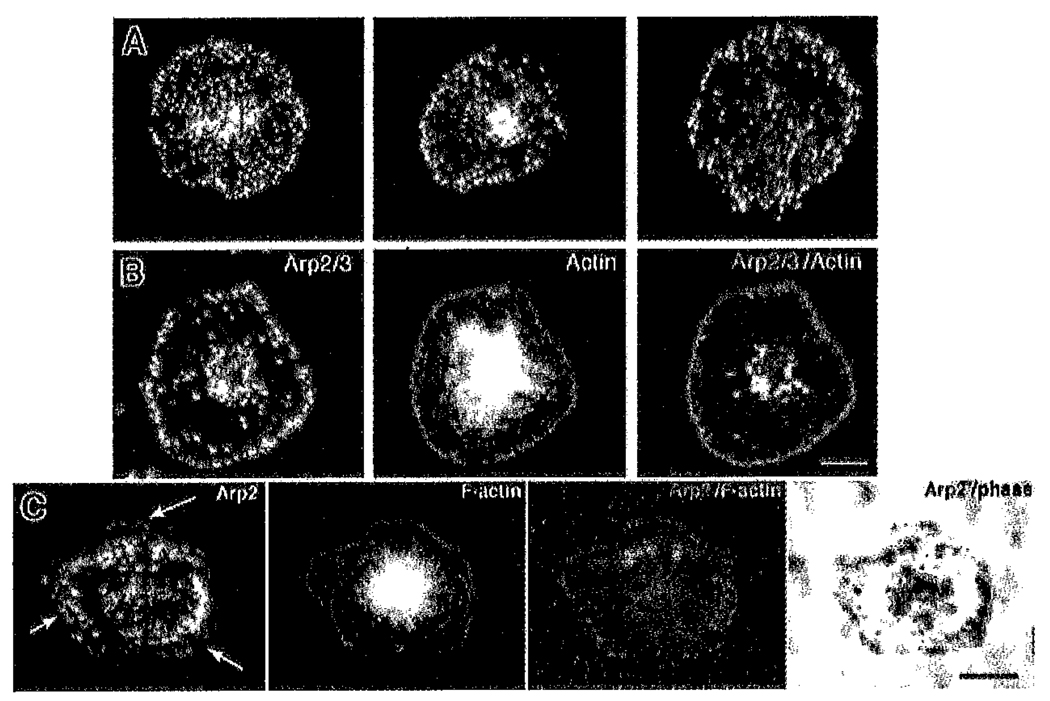
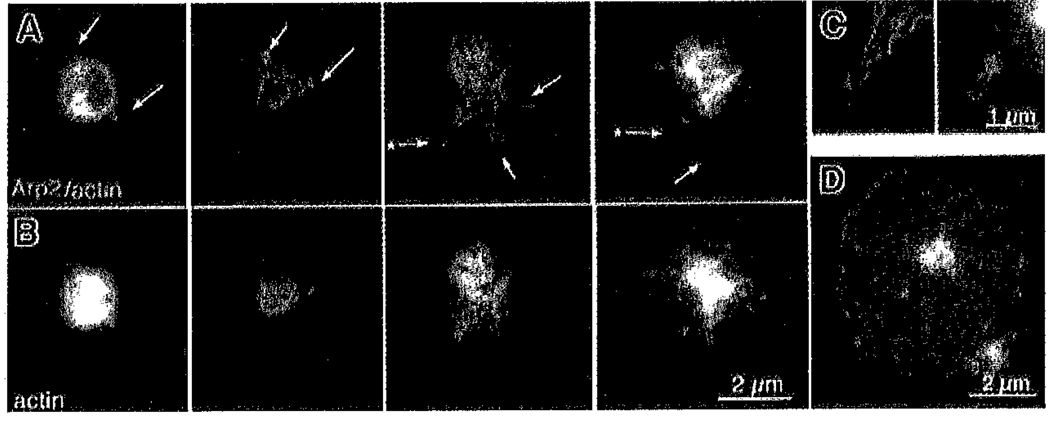
Platelets undergo a series of actin-dependent morphologic changes when activated by thrombin receptor activating peptide (TRAP) or when spreading on glass. Polymerization of actin results in the sequential formation of filopodia, lamellipodia, and stress fibers, but the molecular mechanisms regulating this polymerization are unknown. The Arp2/3 complex nucleates actin polymerization in vitro and could perform this function inside cells as well. To test whether Arp2/3 regulated platelet actin polymerization, we used recombinant Arp2 protein (rArp2) to generate Arp2-specific antibodies (alpha Arp2). Intact and Fab fragments of alpha Arp2 inhibited TRAP-stimulated actin-polymerizing activity in platelet extracts as measured by the pyrene assay. Inhibition was reversed by the addition of rArp2 protein. To test the effect of Arp2/3 inhibition on the formation of specific actin structures, we designed a new method to permeabilize resting platelets while preserving their ability to adhere and to form filopodia and lamellipodia on exposure to glass. Inhibition of Arp2/3 froze platelets at the rounded, early stage of activation, before the formation of filopodia and lamellipodia. By morphometric analysis, the proportion of platelets in the rounded stage rose from 2.85% in untreated to 63% after treatment with alpha Arp2. This effect was also seen with Fab fragments and was reversed by the addition of rArp2 protein. By immunofluorescence of platelets at various stages of spreading, the Arp2/3 complex was found in filopodia and lamellipodia. These results suggest that activation of the Arp2/3 complex at the cortex by TRAP stimulation initiates an explosive polymerization of actin filaments that is required for all subsequent actin-dependent events.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
