

c-Jun N-terminal protein kinase (JNK) 2/3 is specifically activated by stress, mediating c-Jun activation, in the presence of constitutive JNK1 activity in cerebellar neurons
- PMID: 12040039
- PMCID: PMC6758801
- DOI: 10.1523/JNEUROSCI.22-11-04335.2002
c-Jun N-terminal protein kinase (JNK) 2/3 is specifically activated by stress, mediating c-Jun activation, in the presence of constitutive JNK1 activity in cerebellar neurons
Abstract
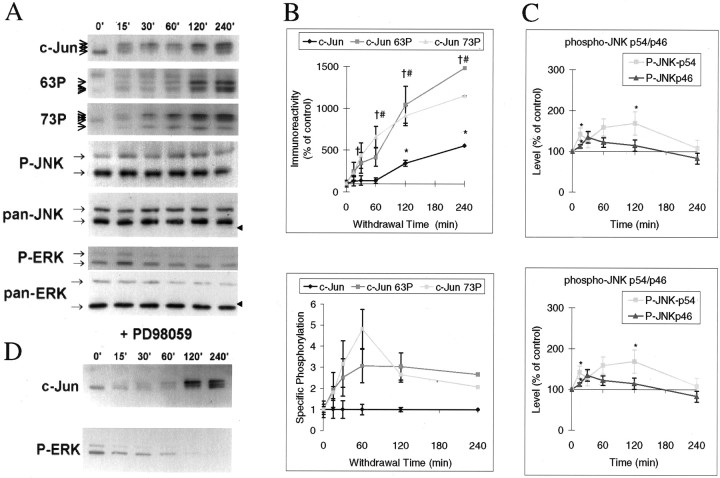
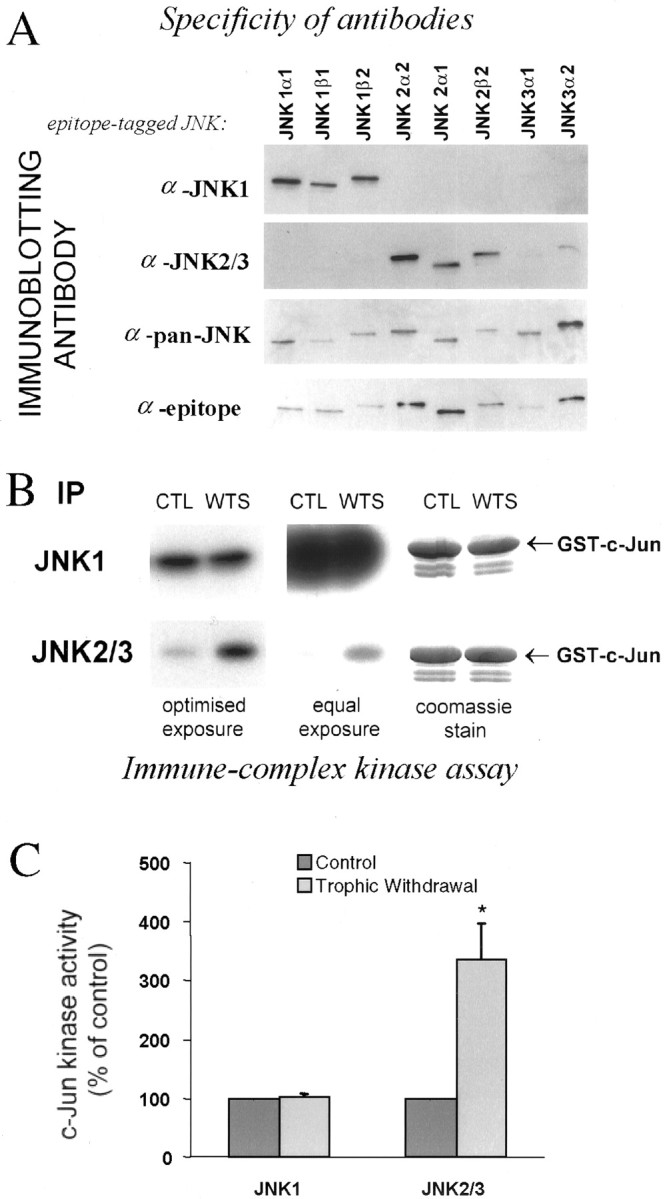
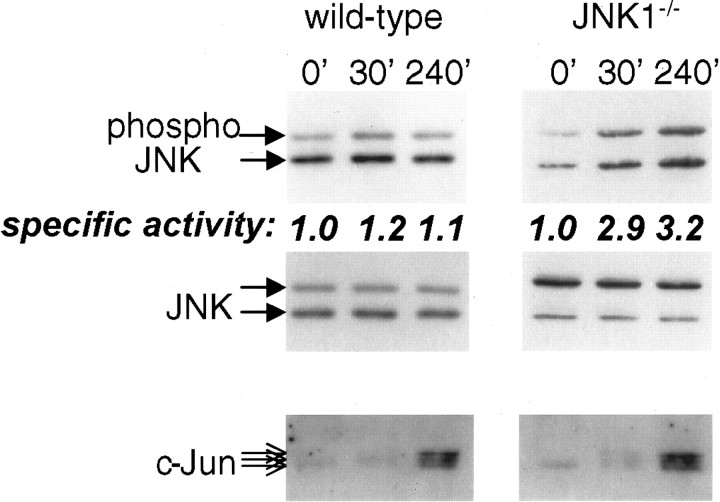
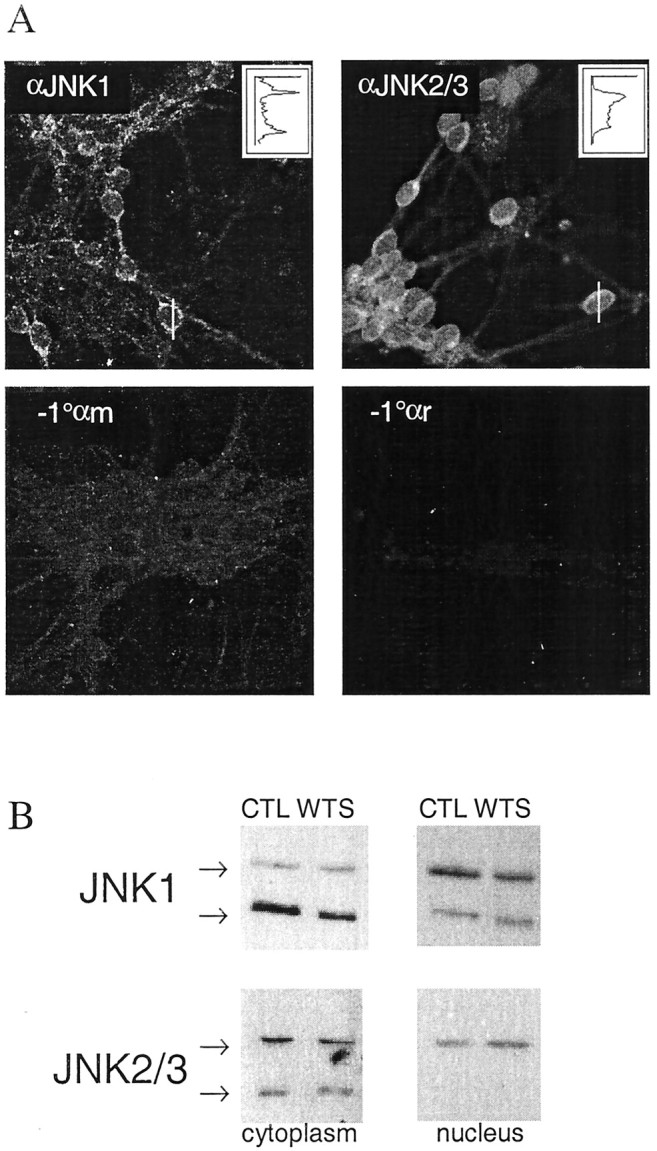
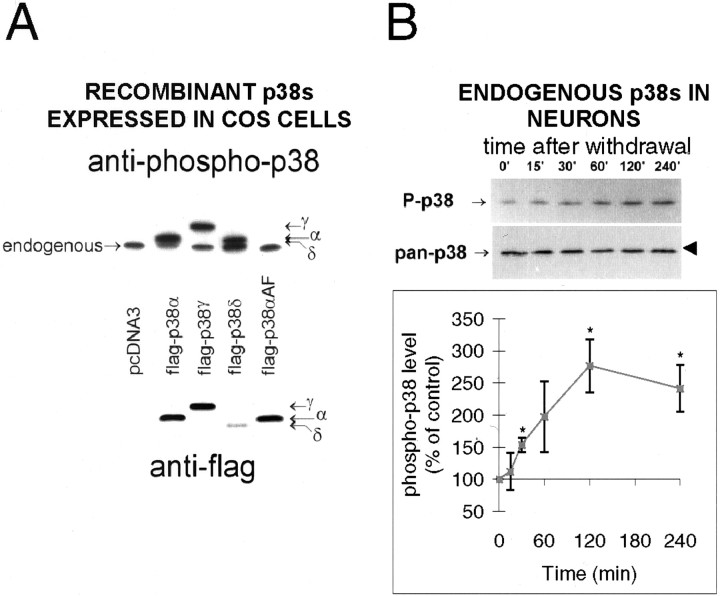
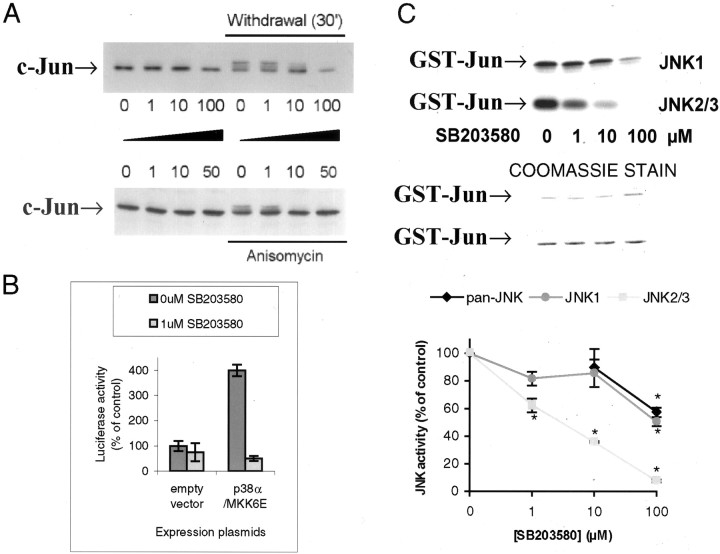
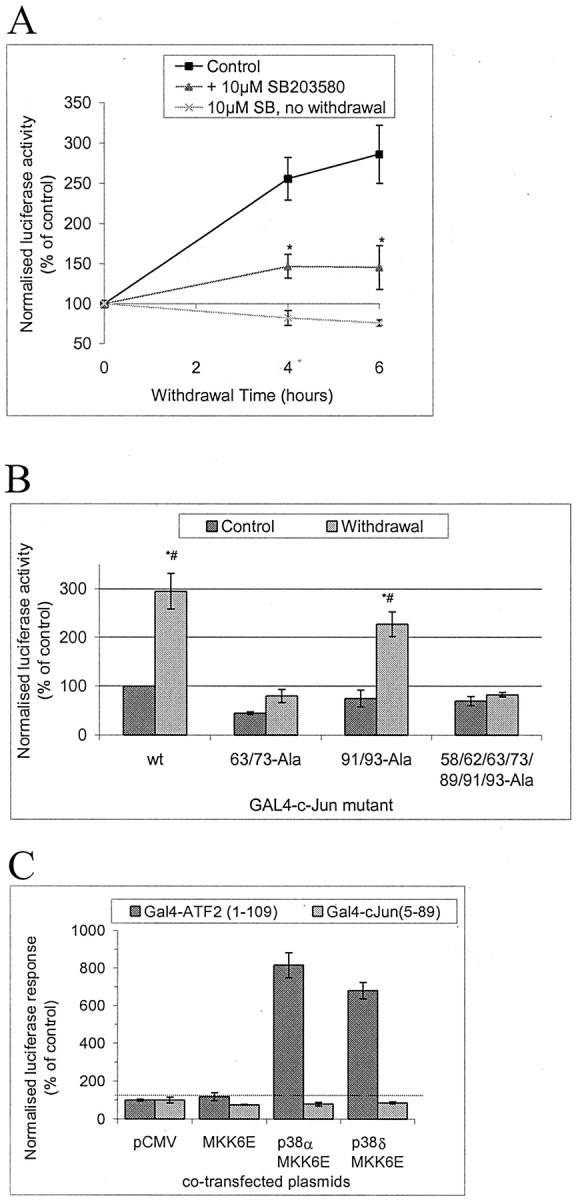
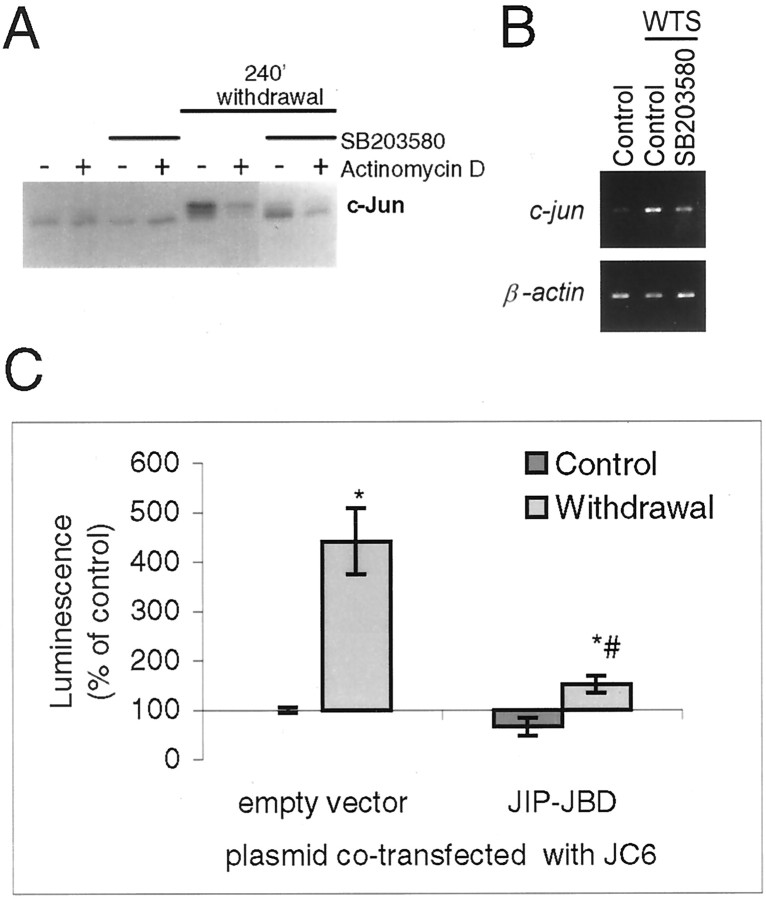
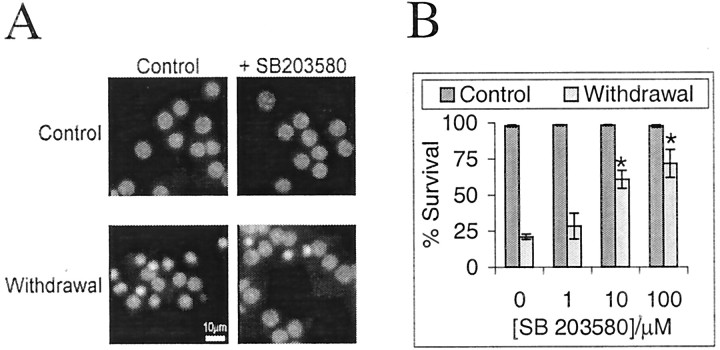
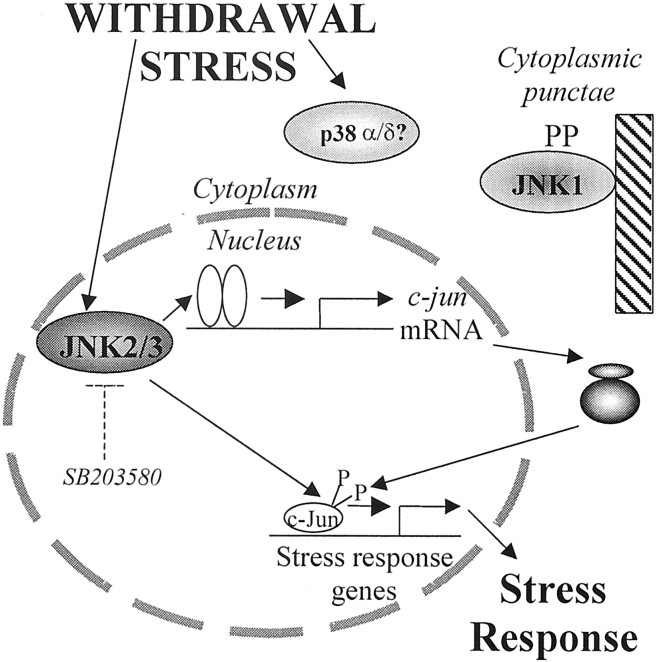
c-Jun is considered a major regulator of both neuronal death and regeneration. Stress in primary cultured CNS neurons induces phosphorylation of c-Jun serines 63 and 73 and increased c-Jun protein. However, total c-Jun N-terminal protein kinase (JNK) activity does not increase, and no satisfactory explanation for this paradox has been available. Here we demonstrate that neuronal stress induces strong activation of JNK2/3 in the presence of constitutively and highly active JNK1. Correspondingly, neurons from JNK1(-/-) mice show lower constitutive activity and considerably higher responsiveness to stress. p38 activity can be completely inhibited without effect on c-Jun phosphorylation, whereas 10 micrometer SB203580 strongly inhibits neuronal JNK2/3, stress-induced c-Jun phosphorylation, induced c-Jun activity, and neuronal death in response to trophic withdrawal stress. Neither constitutive JNK1 activity nor total neuronal JNK activity were significantly affected by this concentration of drug. Thus, neuronal stress selectively activates JNK2/3 in the presence of mechanisms maintaining constitutive JNK1 activity, and this JNK2/3 activity selectively targets c-Jun, which is isolated from constitutive JNK1 activity.
Figures
References
-
- Alessi DR, Cuenda A, Cohen P, Dudley DT, Saltiel AR. PD098059 is a specific inhibitor of the activation of MAPKK in vitro and in vivo. J Biol Chem. 1995;270:27489–27494. - PubMed
-
- Behrens A, Sibilia M, Wagner EF. Amino-terminal phosphorylation of c-Jun regulates stress-induced apoptosis and cellular proliferation. Nat Genet. 1999;21:326–329. - PubMed
-
- Buchdunger E, Zimmermann J, Mett H, Meyer T, Muller M, Druker BJ, Lydon NB. Inhibition of the Abl protein-tyrosine kinase in vitro and in vivo by a 2-phenylaminopyrimidine derivative. Cancer Res. 1996;56:100–104. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous