

Role of p/q-Ca2+ channels in metabotropic glutamate receptor 2/3-dependent presynaptic long-term depression at nucleus accumbens synapses
- PMID: 12040040
- PMCID: PMC6758789
- DOI: 10.1523/JNEUROSCI.22-11-04346.2002
Role of p/q-Ca2+ channels in metabotropic glutamate receptor 2/3-dependent presynaptic long-term depression at nucleus accumbens synapses
Abstract
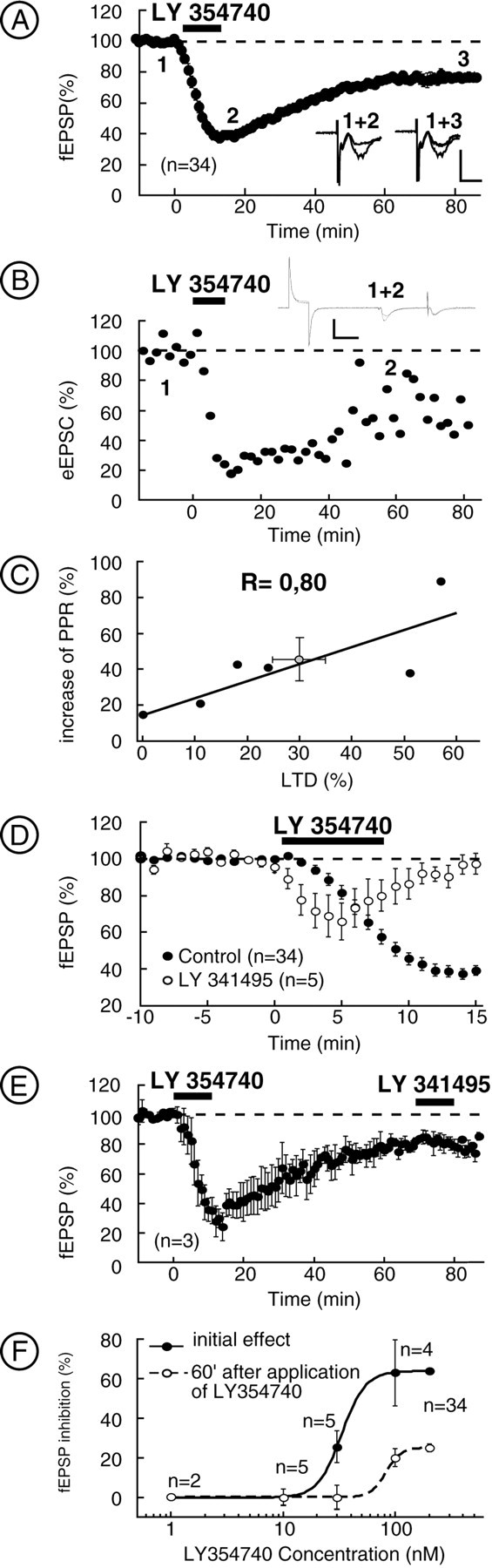
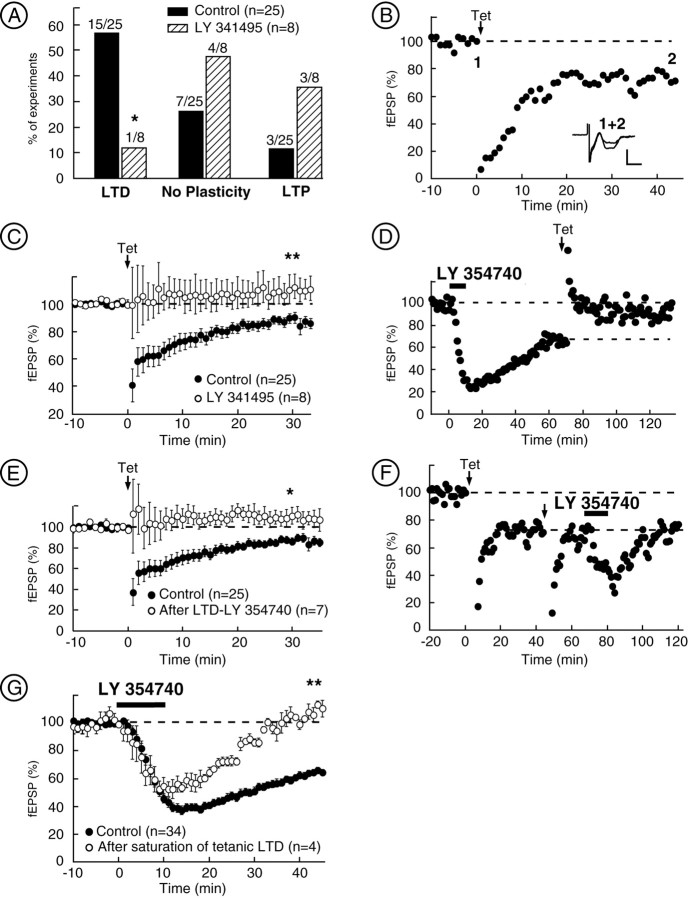
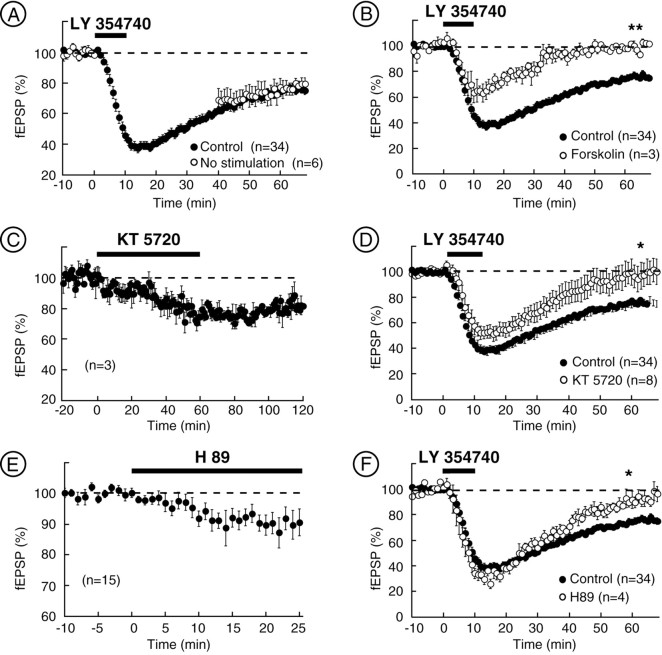
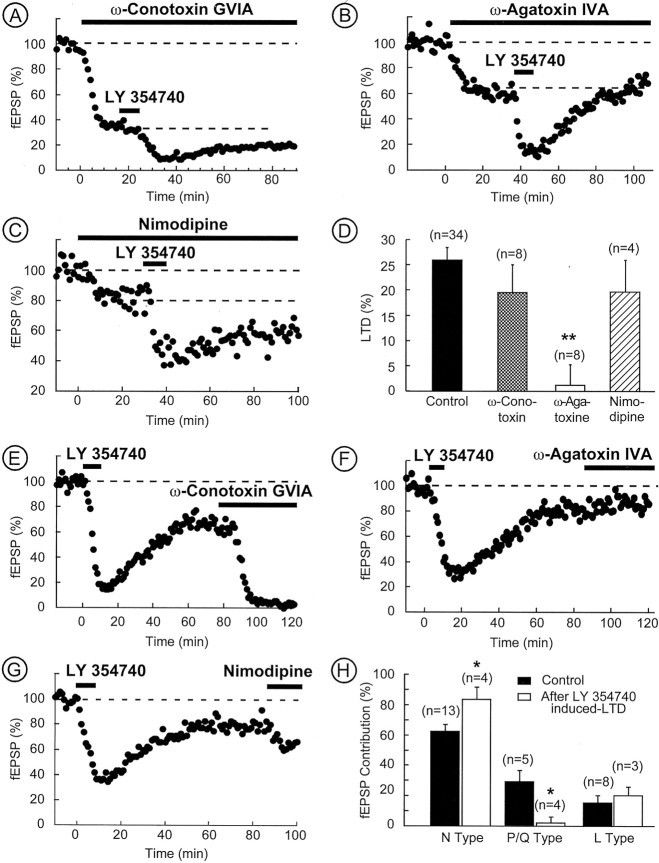
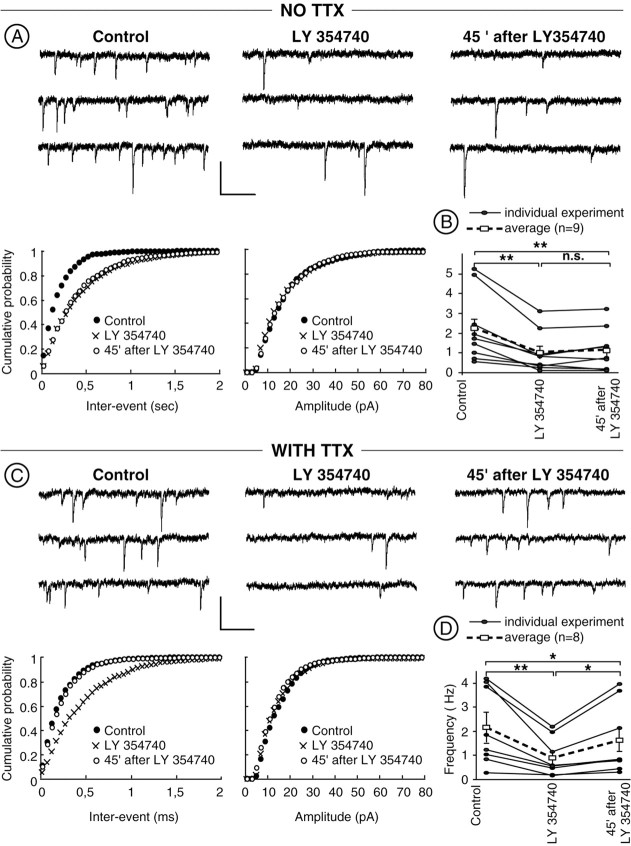
The nucleus accumbens (NAc) is an important cerebral area involved in reward and spatial memory (Pennartz et al., 1994), but little is known about synaptic plasticity in this region. Here, electron microscopy revealed that, in the NAc, metabotropic glutamate receptors 2/3 (mGlu2/3) immunostaining was essentially associated with axonal terminals and glial processes, whereas postsynaptic dendrites and neuronal cell bodies were unstained. Electrophysiological techniques in the NAc slice preparation demonstrated that activation of mGlu2/3 with synaptically released glutamate or specific exogenous agonist, such as LY354740 (200 nm, 10 min), induced long-term depression of excitatory synaptic transmission (mGlu2/3-LTD). Tetanic-LTD and pharmacological mGlu2/3-LTD occluded each other, suggesting common mechanisms. The mGlu2/3-LTD did not require synaptic activity but depended on the cAMP-protein kinase A cascade. Selective inhibition of P/Q-type Ca(2+) channels with omega-agatoxin-IVA occluded the expression of mGlu2/3-LTD, and, conversely, the inhibitory effects of omega-agatoxin-IVA were abolished during mGlu2/3-LTD. Thus, mGlu2/3 play an important role in the control of use-dependent synaptic plasticity at prelimbic cortex-NAc synapses: their activation causes a form of LTD mediated by the long-lasting reduction of P/Q-type Ca(2+)channels contribution to transmitter release.
Figures

References
-
- Anwyl R. Metabotropic glutamate receptors: electrophysiological properties and role in plasticity. Brain Res Brain Res Rev. 1999;29:83–120. - PubMed
-
- Brandon EP, Zhuo M, Huang YY, Qi M, Gerhold KA, Burton KA, Kandel ER, McKnight GS, Idzerda RL. Hippocampal long-term depression and depotentiation are defective in mice carrying a targeted disruption of the gene encoding the RI beta subunit of cAMP-dependent protein kinase. Proc Natl Acad Sci USA. 1995;92:8851–8855. - PMC - PubMed
-
- Cartmell J, Schoepp DD. Regulation of neurotransmitter release by metabotropic glutamate receptors. J Neurochem. 2000;75:889–907. - PubMed
-
- Cartmell J, Monn JA, Schoepp DD. The metabotropic glutamate 2/3 receptor agonists LY354740 and LY379268 selectively attenuate phencyclidine versus d-amphetamine motor behaviors in rats. J Pharmacol Exp Ther. 1999;291:161–170. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous