

Delta 9-tetrahydrocannabinol and cannabinol activate capsaicin-sensitive sensory nerves via a CB1 and CB2 cannabinoid receptor-independent mechanism
- PMID: 12040079
- PMCID: PMC6758782
- DOI: 10.1523/JNEUROSCI.22-11-04720.2002
Delta 9-tetrahydrocannabinol and cannabinol activate capsaicin-sensitive sensory nerves via a CB1 and CB2 cannabinoid receptor-independent mechanism
Abstract
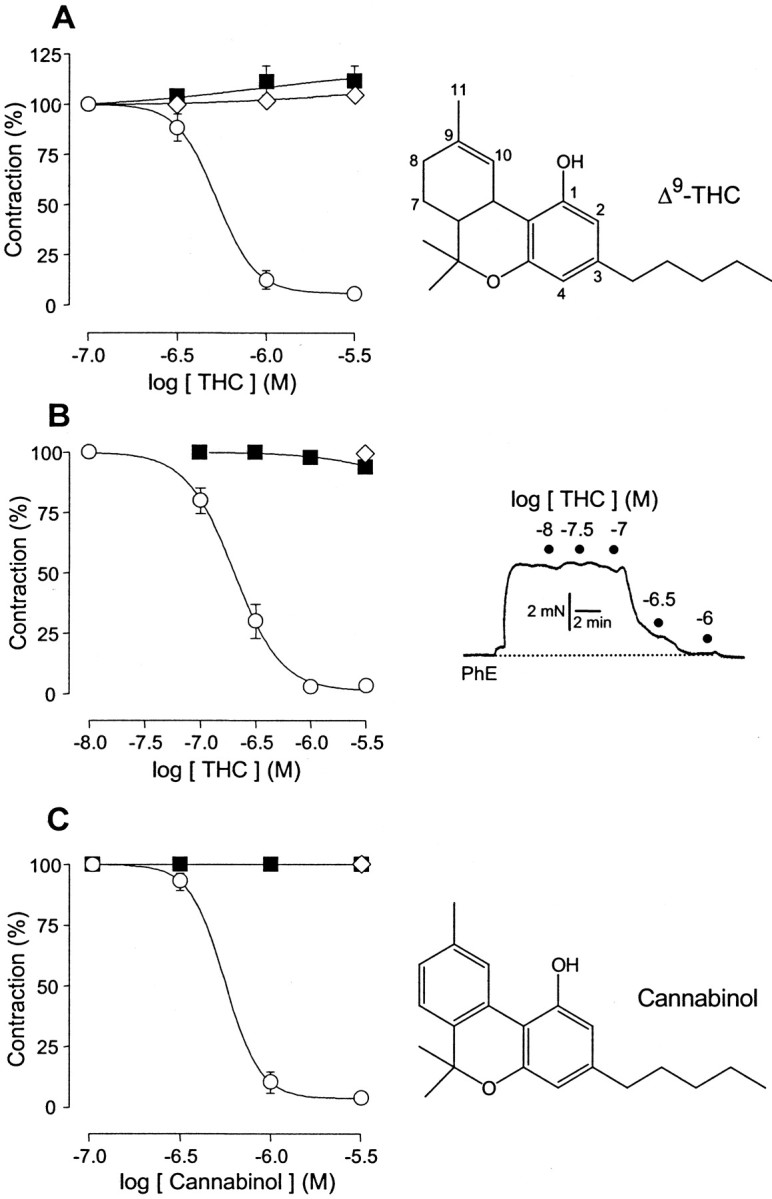
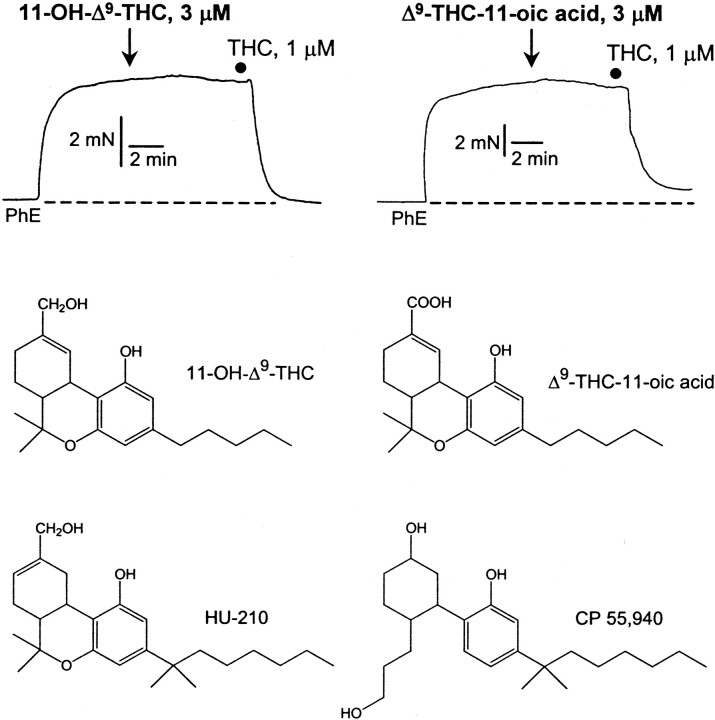
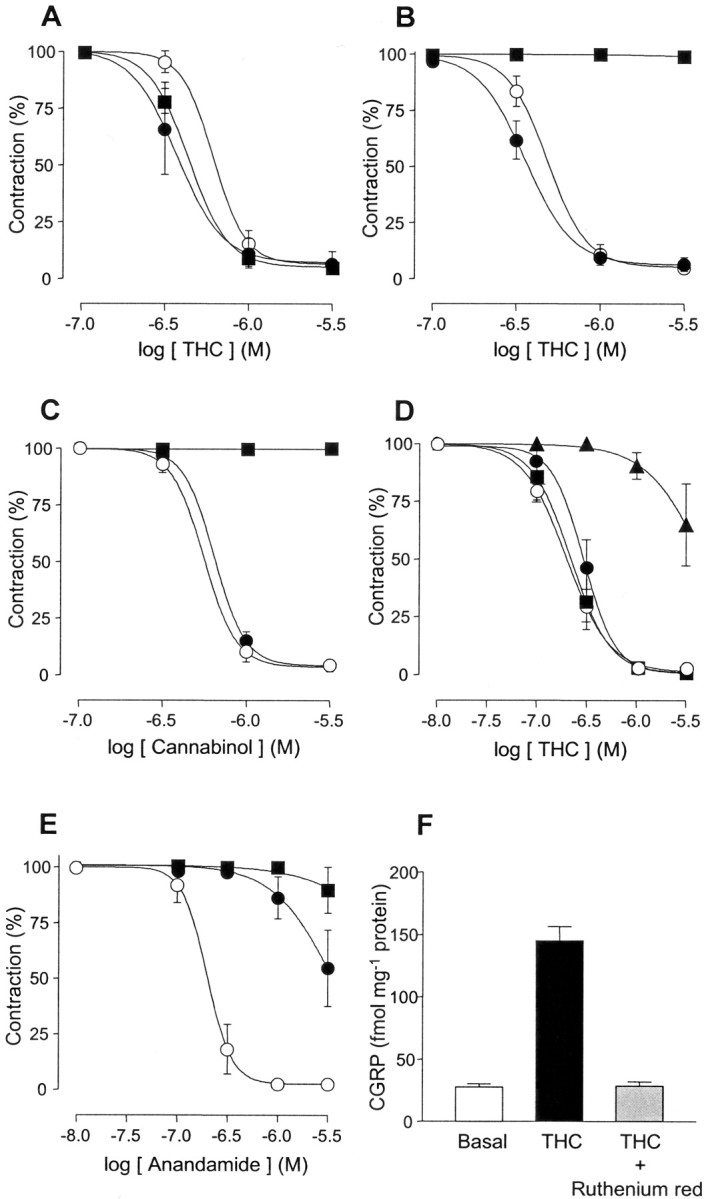
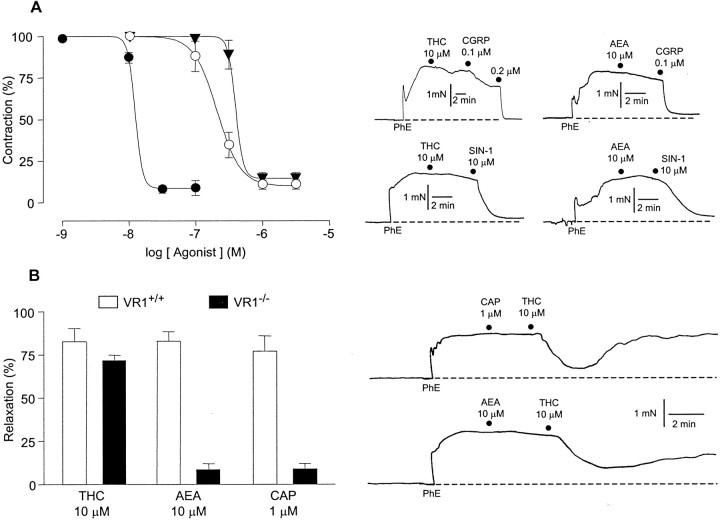
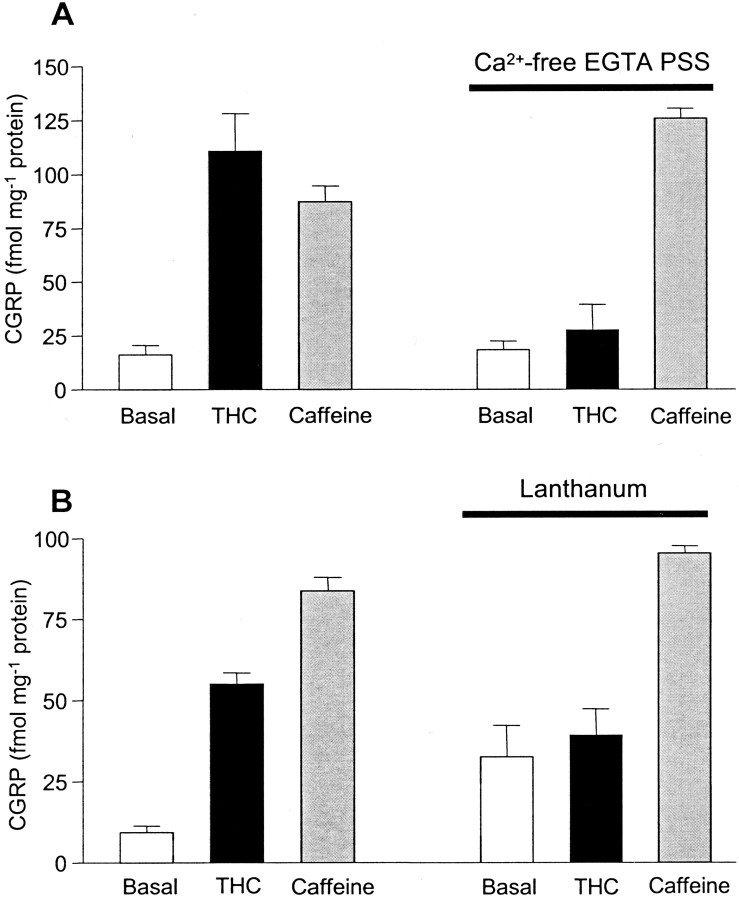
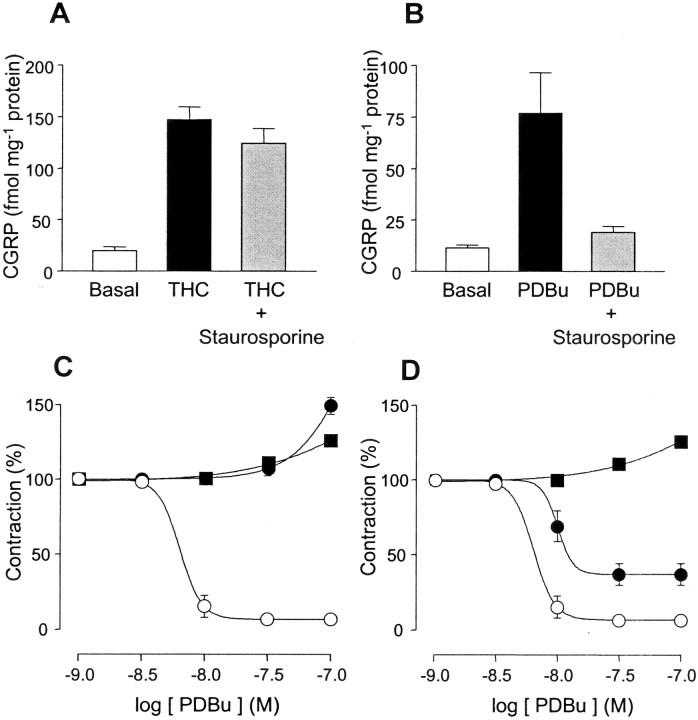
Although Delta(9)-tetrahydrocannabinol (THC) produces analgesia, its effects on nociceptive primary afferents are unknown. These neurons participate not only in pain signaling but also in the local response to tissue injury. Here, we show that THC and cannabinol induce a CB(1)/CB(2) cannabinoid receptor-independent release of calcitonin gene-related peptide from capsaicin-sensitive perivascular sensory nerves. Other psychotropic cannabinoids cannot mimic this action. The vanilloid receptor antagonist ruthenium red abolishes the responses to THC and cannabinol. However, the effect of THC on sensory nerves is intact in vanilloid receptor subtype 1 gene knock-out mice. The THC response depends on extracellular calcium but does not involve known voltage-operated calcium channels, glutamate receptors, or protein kinases A and C. These results may indicate the presence of a novel cannabinoid receptor/ion channel in the pain pathway.
Figures
References
-
- Amann R, Maggi CA. Ruthenium red as a capsaicin antagonist. Life Sci. 1991;49:849–856. - PubMed
-
- Barber LA, Vasko MR. Activation of protein kinase C augments peptide release from rat sensory neurons. J Neurochem. 1996;67:72–80. - PubMed
-
- Bisogno T, Hanus L, De Petrocellis L, Tchilibon S, Ponde DE, Brandi I, Moriello AS, Davis JB, Mechoulam R, Di Marzo V. Molecular targets for cannabidiol and its synthetic analogues: effect on vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide. Br J Pharmacol. 2001;134:845–852. - PMC - PubMed
-
- Blumberg PM. In vitro studies on the mode of action of the phorbol esters, potent tumor promoters: part 1. Crit Rev Toxicol. 1980;8:153–197. - PubMed
-
- Burstein SH. The cannabinoid acids: nonpsychoactive derivatives with therapeutic potential. Pharmacol Ther. 1999;82:87–96. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources