

Energy-efficient neuronal computation via quantal synaptic failures
- PMID: 12040082
- PMCID: PMC6758790
- DOI: 10.1523/JNEUROSCI.22-11-04746.2002
Energy-efficient neuronal computation via quantal synaptic failures
Abstract
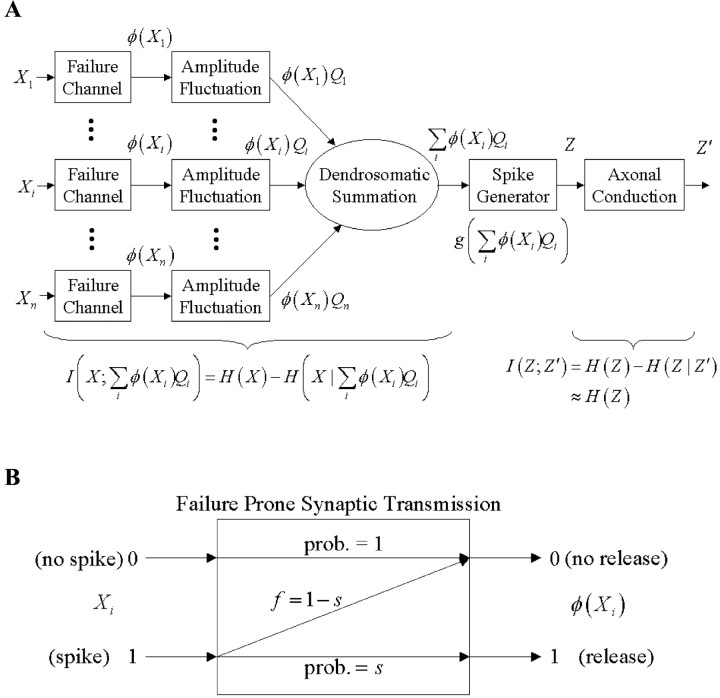
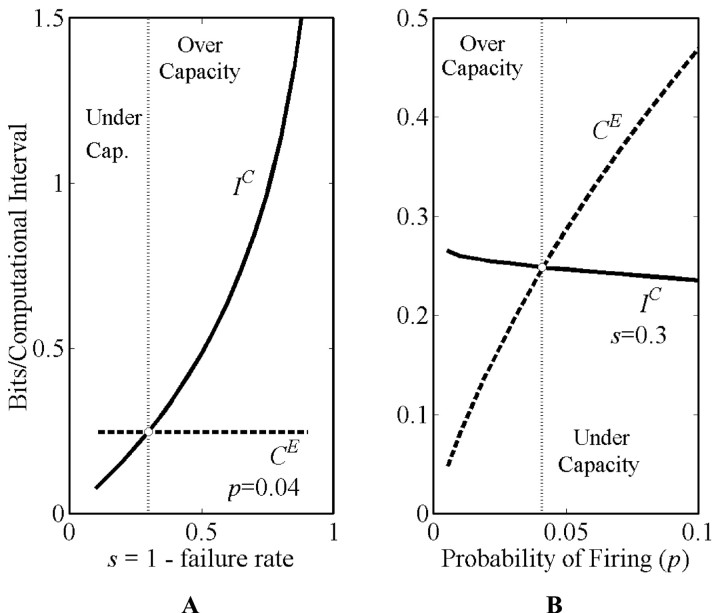
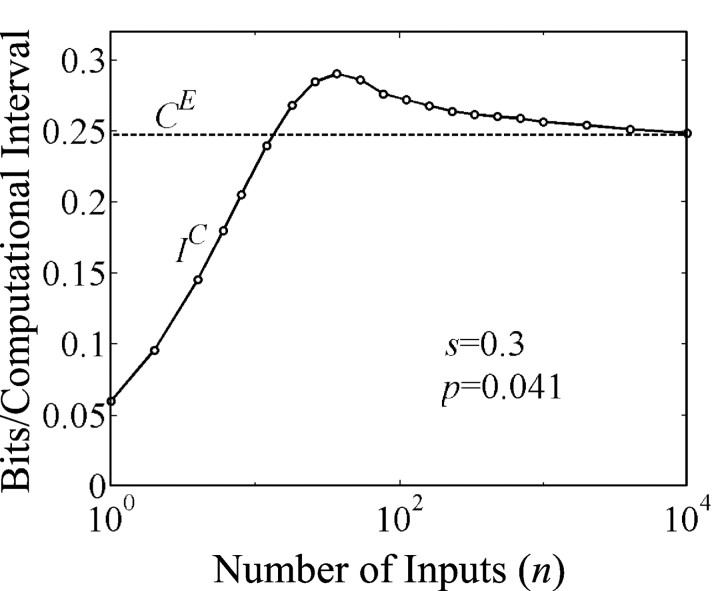
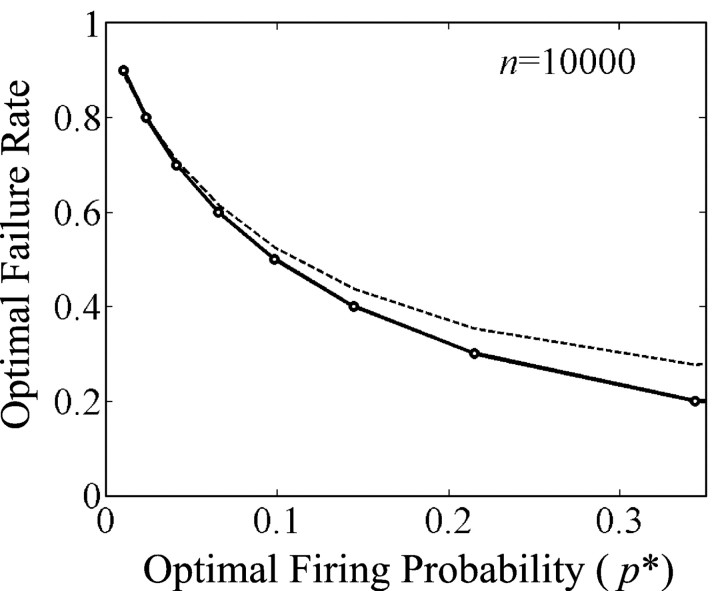
Organisms evolve as compromises, and many of these compromises can be expressed in terms of energy efficiency. For example, a compromise between rate of information processing and the energy consumed might explain certain neurophysiological and neuroanatomical observations (e.g., average firing frequency and number of neurons). Using this perspective reveals that the randomness injected into neural processing by the statistical uncertainty of synaptic transmission optimizes one kind of information processing relative to energy use. A critical hypothesis and insight is that neuronal information processing is appropriately measured, first, by considering dendrosomatic summation as a Shannon-type channel (1948) and, second, by considering such uncertain synaptic transmission as part of the dendrosomatic computation rather than as part of axonal information transmission. Using such a model of neural computation and matching the information gathered by dendritic summation to the axonal information transmitted, H(p*), conditions are defined that guarantee synaptic failures can improve the energetic efficiency of neurons. Further development provides a general expression relating optimal failure rate, f, to average firing rate, p*, and is consistent with physiologically observed values. The expression providing this relationship, f approximately 4(-H(p*)), generalizes across activity levels and is independent of the number of inputs to a neuron.
Figures
References
-
- Abbott L, Sejnowski TJ. Neural codes and distributed representations: foundations of neural computation. MIT; Cambridge, MA: 1999.
-
- Abshire P, Andreou AG. Capacity and energy cost of information in biological and silicon photoreceptors. Proc IEEE. 2001;89:1052–1064.
-
- Andreou AG (1999) Energy and information processing in biological and silicon sensory systems. In: Proceedings of the Seventh International Conference on Microelectronics for Neural, Fuzzy and Bio-Inspired Systems. Los Alamitos, CA, April.
-
- Attwell D, Laughlin SB. An energy budget for signalling in the grey matter of the brain. J Cereb Blood Flow Metab. 2001;21:1133–1145. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous