

Review
doi: 10.1105/tpc.010476.
Gibberellin signaling: biosynthesis, catabolism, and response pathways
Affiliations
- PMID: 12045270
- PMCID: PMC151248
- DOI: 10.1105/tpc.010476
Item in Clipboard
Review
Gibberellin signaling: biosynthesis, catabolism, and response pathways
Plant Cell.
2002.
No abstract available
Figures
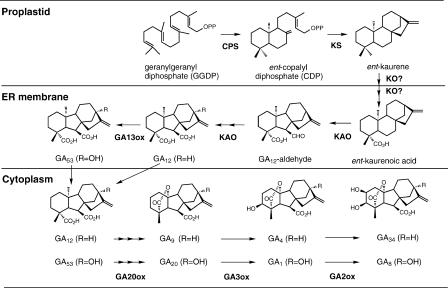
Major GA Biosynthetic and Catabolic Pathways in Higher Plants. The enzyme names are shown in boldface below or to the right of each arrow. GA9 and GA20 also can be converted to GA51 and GA29 by GA2ox. GA4 and GA1 are the bioactive GAs, and GA34 and GA8 are their inactive catabolites.
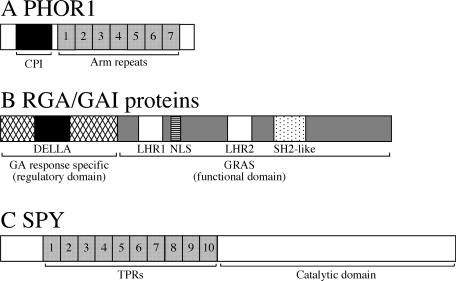
Schemes of PHOR1, RGA/GAI, and SPY. (A) PHOR1 contains a CPI domain, which is a GA-repressible cytosolic retention signal, and the armadillo repeats, which function in nuclear localization and may allow it to interact with transcription factors. (B) RGA/GAI proteins contain a highly conserved C-terminal region that is shared among all GRAS family members. The nuclear localization signal is absent in some of the GRAS members (Pysh et al., 1999; Schumacher et al., 1999; Bolle et al., 2000). RGA/GAI and other nuclear localization signal–containing GRAS family members are likely to function as transcriptional regulators. The N-terminal region of the RGA/GAI proteins is required for the inactivation of these proteins by the GA signal. (C) SPY proteins contain two conserved domains: the TPR domain, which is believed to interact with other proteins; and the catalytic domain, which post-translationally modifies proteins with GlcNAc. Arm, armadillo; LHR, Leu heptad repeat; NLS, nuclear localization signal.

Barley GA Mutants. Plants are of equal age and are (from left to right) Himalaya (wild type), grd2 (GA-deficient putative GA3ox mutant [Chandler and Robertson, 1999]), Sln1d (dominant dwarf), and sln1c (slender).
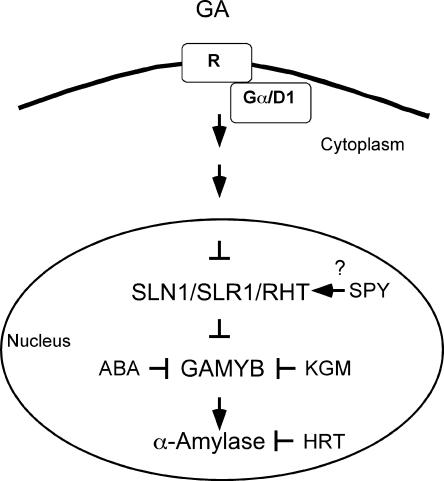
GA Response Pathway Leading to Gene Expression in Cereal Aleurone Cells. ABA, abscisic acid.
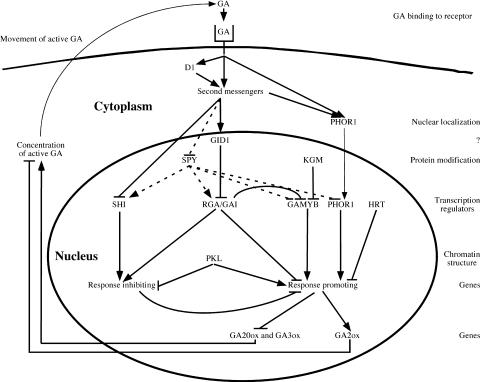
Possible Roles of GA Response Components. The cellular locations and functions of the GA response components discussed in this review are shown. A highly speculative order of action for these components also is shown. Although the diagram shows the components functioning in a single cell, there is no evidence that this occurs. Indeed, some aspects of the response pathway, such as the feedback regulation of GA levels, do not occur in all organs. Bioactive GA binds to an as yet unidentified GA receptor, directly or indirectly activates second messengers and G proteins (D1), and causes PHOR1 to localize to the nucleus and rapid degradation of RGA/GAI. GID1 is a positively acting component that is required for the degradation of GAI/RGA proteins. Although GID1 is shown in the cytosol, the cellular location has not been determined. In the nucleus, SPY and KGM inhibit GA responses by GlcNAc, modifying or phosphorylating component(s) of the pathway. Several putative transcription regulators (SHI, RGA/GAI proteins, GAMYB, PHOR1, and HRT) modify the transcription of genes encoding products that either promote or inhibit the GA response. Some of the products of these genes may be positive or negative regulators acting in a transcriptional cascade that leads to the GA response. The maintenance of chromatin structure by PKL is important for the proper expression of genes involved in the response. One outcome of the GA response is reduced transcription of GA20ox and GA3ox and enhanced expression of GA2ox, which leads to a reduction in the pool of bioactive GA.
References
-
- Aach, H., Bode, H., Robinson, D.G., and Graebe, J.E. (1997). ent-Kaurene synthase is located in proplastids of meristematic shoot tissues. Planta 202 211–219.
-
- Amador, V., Monte, E., García-Martínez, J.-L., and Prat, S. (2001). Gibberellins signal nuclear import of PHOR1, a photoperiod-responsive protein with homology to Drosophila armadillo. Cell 106 343–354. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
