

Review
doi: 10.1105/tpc.001768.
Ethylene biosynthesis and signaling networks
Affiliations
- PMID: 12045274
- PMCID: PMC151252
- DOI: 10.1105/tpc.001768
Item in Clipboard
Review
Ethylene biosynthesis and signaling networks
Plant Cell.
2002.
No abstract available
Figures
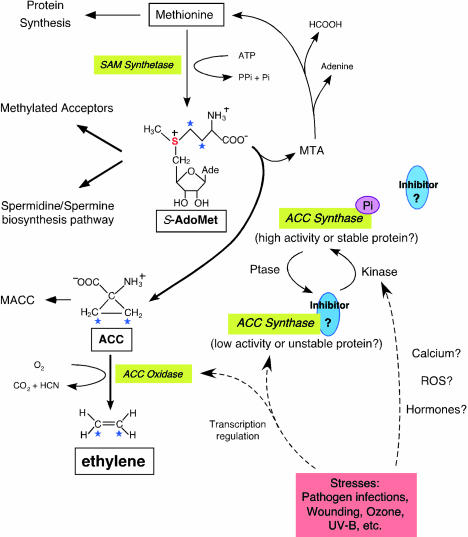
Biosynthetic Pathway and Regulation of Ethylene. The formation of S-AdoMet is catalyzed by SAM synthetase from the methionine at the expense of one molecule of ATP per molecule of S-AdoMet synthesized. S-AdoMet is the methyl group donor for many cellular molecules (Methylated Acceptors), including nucleic acids, proteins, and lipids. In addition, S-AdoMet is the precursor of the polyamine synthesis pathway (Spermidine/Spermine biosynthesis pathway). ACC is the immediate precursor of ethylene. The rate-limiting step of ethylene synthesis is the conversion of S-AdoMet to ACC by ACC synthase under most conditions. MTA is the by-product generated along with ACC production by ACC synthase. Recycling of MTA back to methionine conserves the methylthio group and is able to maintain a constant concentration of cellular methionine even when ethylene is rapidly synthesized. Malonylation of ACC to malonyl-ACC (MACC) deprives the ACC pool and reduces the ethylene production. ACC oxidase catalyses the final step of ethylene synthesis using ACC as substrate and generates carbon dioxide and cyanide. Transcriptional regulation of both ACC synthase and ACC oxidase is indicated by dashed arrows. Reversible phosphorylation of ACC synthase is hypothesized and may be induced by unknown phosphatases (Ptase) and kinases, the latter presumably activated by stresses. Both native and phosphorylated form (ACC synthase-Pi) of ACC synthase are functional, although the native ACC synthase may be less stable or active in vivo. A hypothetical inhibitor is associated with ACC synthase at the carboxyl end and may be dissociated from the enzyme if it is modified by phosphorylation at the vicinity.
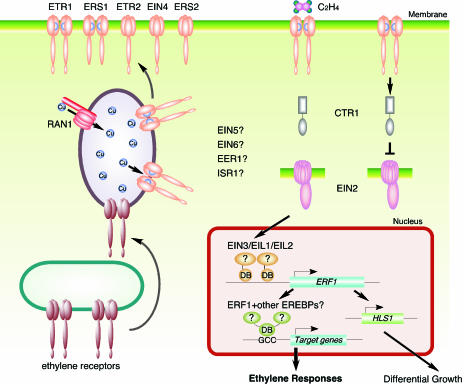
Model of the Ethylene Signal Transduction Pathway. There are five ethylene receptors in Arabidopsis, ETR1, ETR2, EIN4, ERS1, and ERS2. ETR1 and ERS1 contain three transmembrane domains and a conserved histidine kinase domain, and have been shown to function as homodimers. ETR2, EIN4, and ERS2 have four membrane-spanning regions and a degenerate histidine kinase domain. Only ETR1, ETR2, and EIN4 have receiver domains at their C termini. Ethylene binding occurs at the N-terminal transmembrane domain of the receptors, and a copper co-factor is required for the binding. RAN1, a copper transporter, is involved in delivery of copper to the ethylene receptor. In the absence of an ethylene signal, ethylene receptors activate a Raf-like kinase, CTR1, and CTR1 in turn negatively regulates the downstream ethylene response pathway, possibly through a MAP-kinase cascade. Binding of ethylene inactivates the receptors, resulting in deactivation of CTR1, which allows EIN2 to function as a positive regulator of the ethylene pathway. EIN2 contains the N-terminal hydrophobic domain similar to the Nramp metal transporter proteins and the novel hydrophilic C terminus. EIN2 positively signals downstream to the EIN3 family of transcription factors located in the nucleus. EIN3 binds to the promoter of ERF1 gene and activates its transcription in an ethylene-dependent manner. Transcription factors ERF1 and other EREBPs can interact with the GCC box in the promoter of target genes and activate downstream ethylene responses.
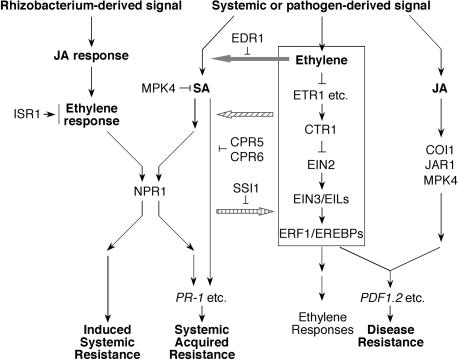
Interactions between the Ethylene Signal Transduction Pathway and Plant Disease Resistance. The ethylene signal transduction pathway can interact with the JA pathway to co-regulate expression of a subset of defense-related PR genes, for example, PDF1.2, involved in plant disease resistance. Meanwhile, there are considerable interactions between JA/ethylene- and SA-dependent pathways in systemic acquired resistance. In edr1 mutant, ethylene potentiates SA-mediated PR-1 gene expression. In the absence of CPR5 and CPR6, the ethylene pathway can also activate SA-dependent PR-1 gene expression independent of NPR1 to promote systemic acquired resistance. In the ssi1 mutant, the JA/ethylene-dependent PDF1.2 gene is constitutively expressed. Moreover, the ethylene pathway is also required for the rhizobacteria-mediated induced systemic resistance, which is independent of SA and pathogenesis-related gene activation. Ethylene signaling acts downstream of the JA pathway but upstream of NPR1 in ISR activation. Plants that lack ISR1 fail to develop ISR and display ethylene insensitivity. Arrows indicate positive regulation, and open blocks indicate negative regulation.
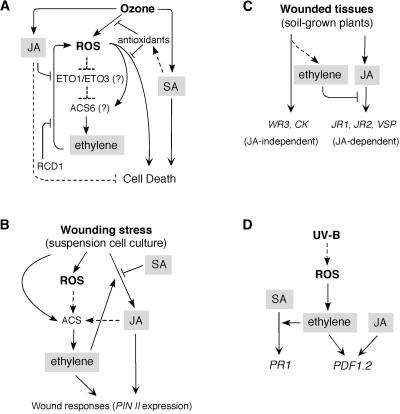
Interactions among Ethylene, JA, and SA in Abiotic Stresses. (A) Ozone stress. (B) Wound response in tomato suspension cell culture. (C) Wounded tissues (local responses). (D) UV-B stress. ACS, ACC synthase; ETO1 (ETO3), ethylene overproducer; RCD1, radical-induced cell death. Arrows indicate positive regulation, and open blocks indicate negative regulation. Dashed lines indicate possible or indirect interactions.
References
-
- Abel, S., Nguyen, M.D., Chow, W., and Theologis, A. (1995). ACS4, a primary indoleacetic acid-responsive gene encoding 1-aminocyclopropane-1-carboxylate synthase in Arabidopsis thaliana. Structural characterization, expression in Escherichia coli, and expression characteristics in response to auxin. J. Biol. Chem. 270 19093–19099. - PubMed
-
- Alonso, J.M., Hirayama, T., Roman, G., Nourizadeh, S., and Ecker, J.R. (1999). EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science 284 2148–2152. - PubMed
-
- Arteca, J.M., and Arteca, R.N. (1999). A multi-responsive gene encoding 1-aminocyclopropane-1-carboxylate synthase (ACS6) in mature Arabidopsis leaves. Plant Mol. Biol. 39 209–219. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
