

Review
doi: 10.1105/tpc.000596.
Cell signaling during cold, drought, and salt stress
Affiliations
- PMID: 12045276
- PMCID: PMC151254
- DOI: 10.1105/tpc.000596
Item in Clipboard
Review
Cell signaling during cold, drought, and salt stress
Plant Cell.
2002.
No abstract available
Figures
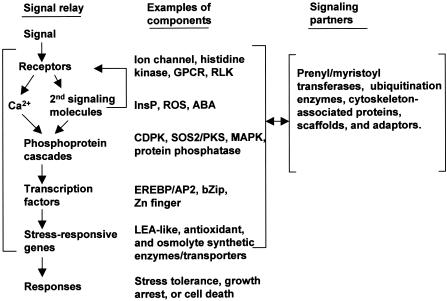
A Generic Pathway for the Transduction of Cold, Drought, and Salt Stress Signals in Plants. Examples of signaling components in each of the steps are shown (for more detailed information, see Xiong and Zhu, 2001). Secondary signaling molecules can cause receptor-mediated Ca2+ release (indicated with a feedback arrow). Examples of signaling partners that modulate the main pathway are also shown. These partners can be regulated by the main pathway. Signaling can also bypass Ca2+ or secondary signaling molecules in early signaling steps. GPCR, G-protein coupled receptor; InsP, inositol polyphosphates; RLK, receptor-like kinase. Other abbreviations are given in the text.
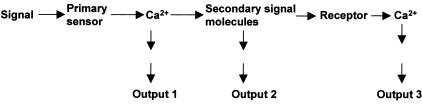
Repetitive Ca2+ Transients upon the Perception of a Primary Signal. The primary increase in cytosolic Ca2+ facilitates the generation of secondary signaling molecules, which stimulate a second round of transient Ca2+ increases, both locally and globally. These second Ca2+ transients may feedback regulate each of the previous steps (not shown). Ca2+ transients from different sources may have different biological significance and result in different outputs, as shown. Secondary signaling molecules such as ROS can also directly regulate signal transduction without Ca2+ (Output 2).
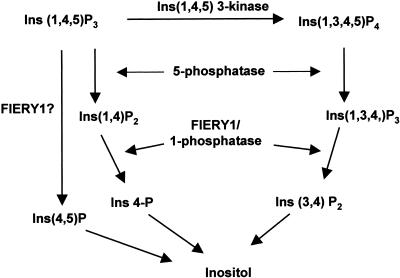
Potential Pathways for Inositol 1,4,5-Trisphosphate (IP3) Degradation in Plants. The pathways are drawn on the basis of information from animal systems. FIERY1 inositol polyphosphate 1-phosphatase can hydrolyze Ins(1,4)P2 and Ins(1,3,4)P3. A potential pathway mediated by FIERY1 with direct hydrolysis of IP3 at the 1-position is also indicated (with a question mark). 5-phosphatase, inositol polyphosphate 5-phosphatase.
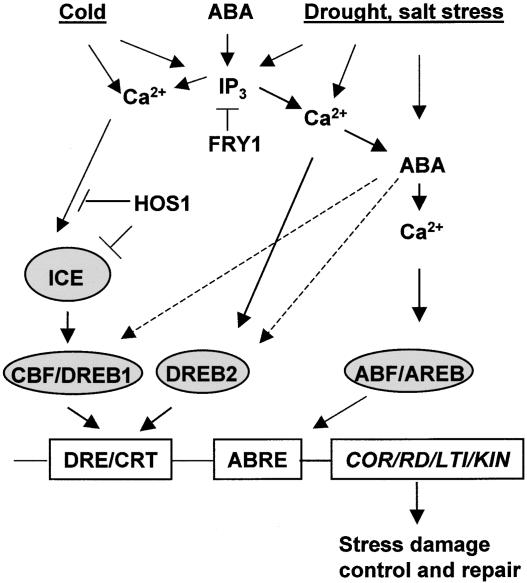
Pathways for the Activation of the LEA-Like Class of Stress-Responsive Genes with DRE/CRT and ABRE cis Elements. Cold, drought, salt stress, and ABA can activate these genes through stress-inducible transcription factors CBF/DREB1 and DREB2, and ABA-inducible bZIP transcription factors ABF/AREB (Shinozaki and Yamaguchi-Shinozaki, 2000). An unidentified transcriptional activator, ICE (inducer of CBF expression) (Thomashow, 2001), is indicated. IP3 is involved in the signaling, as revealed by genetic identification of the FRY1 locus, which negatively regulates IP3 levels and stress signaling (Xiong et al., 2001c). The HOS1 locus negatively regulates cold signaling, presumably by targeting ICE or upstream signaling components for degradation (Lee et al., 2001). DREB2-mediated gene activation also depends on ABA-dependent posttranscriptional/translational modifications of CBF/DREB1 or DREB2 or associated coactivators (indicated with dashed arrows). COR, cold regulated; KIN, cold induced; LTI, low-temperature induced; RD, responsive to dehydration.
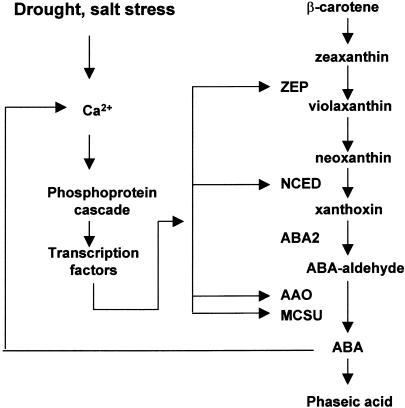
Pathway and Regulation of ABA Biosynthesis. ABA is synthesized from a C40 precursor β-carotene via the oxidative cleavage of neoxanthin and a two-step conversion of xanthoxin to ABA via ABA-aldehyde. Environmental stress such as drought, salt and, to a lesser extent, cold stimulates the biosynthesis and accumulation of ABA by activating genes coding for ABA biosynthetic enzymes. Stress activation of ABA biosynthetic genes is probably mediated by a Ca2+-dependent phosphorelay cascade, as shown at left. In addition, ABA can feedback stimulate the expression of ABA biosynthetic genes, also likely through a Ca2+-dependent phosphoprotein cascade (Xiong et al., 2001a, ; L. Xiong and J.K. Zhu, unpublished data). Also indicated is the breakdown of ABA to phaseic acid. AAO, ABA-aldehyde oxidase; MCSU, molybdenum cofactor sulfurase; NCED, 9-cis-epoxycarotenoid dioxygenase; ZEP, zeaxanthin epoxidase.
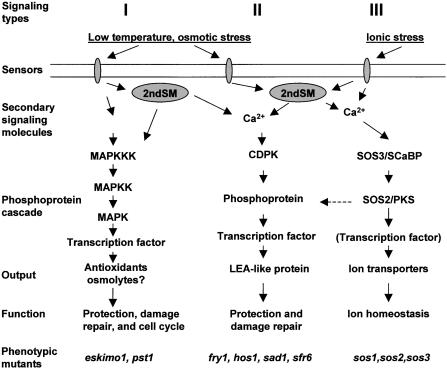
Major Types of Signaling for Plants during Cold, Drought, and Salt Stress. Representative cascades, outputs, biological functions, and examples of mutants with phenotypes indicative of defects in the respective biological functions are shown. Type I signaling involves the generation of ROS scavenging enzymes and antioxidant compounds as well as osmolytes. The involvement of a MAPK pathway in the production of osmolytes in plants has not been demonstrated experimentally. Under osmotic stress, altered MAPK signaling may contribute to changed cell cycle regulation and growth retardation. Type II signaling involves the production of stress-responsive proteins mostly of undefined functions. Pathways within Type II signaling are shown in Figure 4. Type III signaling involves the SOS pathway which is specific to ionic stress. Signaling events for homologs of SOS3 (SCaBP) and SO2 (PKS) are tentatively grouped with SOS3 and SOS2, yet these SCaBP-PKS pathways are not necessarily related to ion homeostasis. Connections between different types of signaling events are indicated with dashed lines. Arrows indicate the direction of signal flux. Primary sensors are shown to be localized in the membrane. Receptors for secondary signaling molecules (2ndSM) are not shown.
References
-
- Allen, G.J., Chu, S.P., Harrington, C.L., Schumacher, K., Hoffman, T., Tang, Y.Y., Grill, E., and Schroeder, J.I. (2001). A defined range of guard cell calcium oscillation parameters encodes stomatal movements. Nature 411 1053–1057. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
