

Review
doi: 10.1105/tpc.001065.
Small GTPases: versatile signaling switches in plants
Affiliations
- PMID: 12045289
- PMCID: PMC151267
- DOI: 10.1105/tpc.001065
Item in Clipboard
Review
Small GTPases: versatile signaling switches in plants
Plant Cell.
2002.
No abstract available
Figures
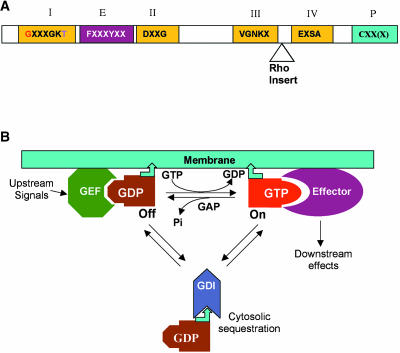
Conserved Structure and Regulation of Small GTPases. (A) Conserved structure of small GTPase. All small GTPases contain four conserved domains for guanine nucleotide binding and GTPase activities (I through IV) and an effector domain (E). Residues in red have been used for the generation of CA mutants, and those in purple for DN mutants (see Figure 2). The Rho insert (10 to 12 amino acids) is only found in Rho GTPases. Rab, Ras, and Rho GTPases also contain a C-terminal motif for prenylation (P). The motif can be CAAL (C, cysteine; A, aliphatic amino acid; L, leucine) for geranylgeranylation by geranylgeranyltransferase I (GGTase I), CC/CXC for geranylgeranylation by GGTase II, or CAAX (X indicates any amino acid except for leucine and phenyalanine) for farnesylation. Arf is myristoylated at the N terminus, but Ran has no known modification. (B) A general scheme for the regulation and action of small GTPases. GAP, GTPase-activating protein; GDI, guanine nucleotide dissociation inhibitor; GEF, guanine nucleotide exchange factor. Bent arrowhead indicates a lipid moiety that becomes attached to membranes.
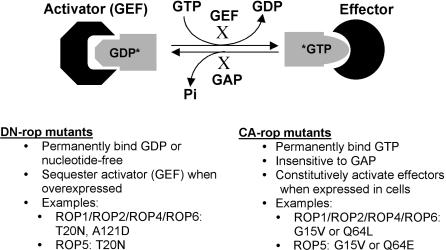
Schematics of CA and DN rop Mutants Used for Functional Analysis. The DN mutant of GTPase is locked in GDP-bound or nucleotide-free form, and the CA mutant is locked in GTP-bound form. Overexpression of DN mutants blocks Rop activation by sequestering activators, whereas CA mutants are insensitive to the action of RopGAPs and thus permanently activate effectors. Examples shown are those used for the investigation of the function of Arabidopsis ROPs (Kost et al., 1999; Li et al., 1999; Lemichez et al., 2001; Li et al., 2001). X indicates the inhibition of the cycling between GDP and GTP forms by the dominant mutations.
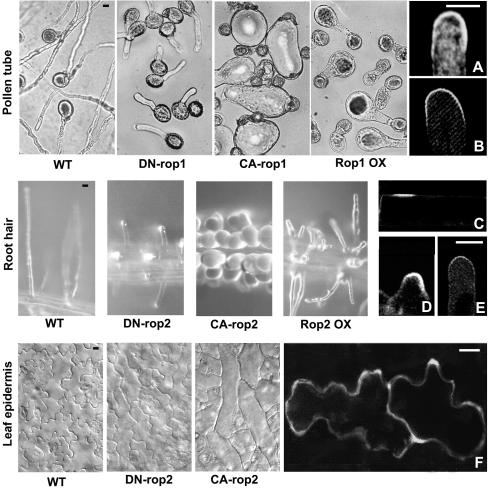
Rop Regulates Cell Polarity Development and Polar Growth through Recruitment to and Activation at the Site of Action. Typical cell-shape phenotypes induced by CA-rop, DN-rop, or Rop OX in different cells (Li et al., 1999; Fu et al., 2002; Jones et al., 2002) are shown at left, and Rop localization or its activation is shown at right. (A) Rop localization to the apical PM region in pea pollen tubes as shown by indirect immunoflurorescence (Lin et al., 1996). (B) The localization of GFP-tagged RIC1 in tobacco pollen tubes, indicating the localization of GTP-bound active Rop to the apical PM region (Wu et al., 2001). (C) to (E) The localization of GFP-tagged ROP2 to the future hair forming site in the epidermal cell (C), the tip of swelling prior to the initiation of tip growth (D), and the tip of growing root hair (E) (Jones et al., 2002). (F) Preferential localization of GFP-ROP2 to the site of lobe formation in expanding leaf epidermal cells (Fu et al., 2002). All images shown were obtained using confocal microscopy. Scale bars = 10 μm.
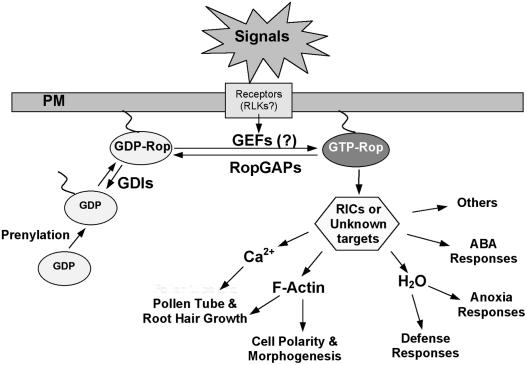
A Generalized Scheme Illustrating the Functional Diversity of Rop GTPases. Rop signaling controls many processes and involves many Rop-interacting proteins. Different Rops may interact differentially with different Rop regulators and target proteins to produce functionally distinct Rop signaling pathways. Rop regulation involves not only activation by an unknown mechanism and deactivation by RopGAPs and Rop GDIs, but also compartmentation in specific membranes or membrane domains. In addition to the PM as shown in this figure, Rop is also found in specific PM domains and other compartments such as vacuoles. Targeting to specific compartments could require specific unknown recruiting or docking proteins. Functionally distinct Rop targets including various RICs and possibly other unknown proteins are proposed to control specific downstream effects to achieve functional specificity of each Rop-dependent pathway.
References
-
- Arabidopsis Genome Initiative. (2000). Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408 796–815. - PubMed
-
- Aspenstrom, P. (1999). Effectors of the Rho GTPases. Curr. Opin. Cell Biol. 11 95–102. - PubMed
-
- Baluska, F., Salaj, J., Mathur, J., Braun, M., Jasper, F., Samaj, J., Chua, N.H., Barlow, P.W., and Volkmann, D. (2000). Root hair formation: F-actin-dependent tip growth is initiated by local assembly of profilin-supported F-actin meshworks accumulated within expansin-enriched bulges. Dev. Biol. 227 618–632. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
